The high subarctic seas (Norwegian/Barents Sea, Labrador Sea, and Bering Sea) are characterized by high biological productivity and the seasonal presence of sea ice (Hunt and Drinkwater, 2005). Differences in physiography and orientation or exposure to the dominant forcing mechanisms help to determine the landscape ecology of these areas (Bailey et al., 2003; Ciannelli and Bailey, 2005; Stabeno et al., 2006). In the last decade, global temperatures have reached some of the highest levels ever recorded and projections of future (after 2030) temperature suggest that the greatest rates of change will be at high latitudes (IPCC, 2007). Increases in global temperatures have begun to affect the areal extent, concentration, and thickness of ice in both the polar regions and in the subarctic seas (IPCC, 2007). It is imperative that we understand the role played by physical mechanisms in arctic ecosystems, including how sea ice impacts the structure, function, and productivity of these ecosystems. The focus of this article is an examination of the physics, chemistry, chlorophyll, and zooplankton over the eastern Bering Sea shelf from spring through summer 2005. Since 2005 was a particularly warm year in the Bering Sea, it may provide insight into the future of the Bering Sea ecosystem.
Sea ice is a critical component of the oceanography of the Bering Sea shelf. During winter, frigid winds blow southward over the eastern Bering Sea, forming ice in the lee of headlands and islands. In these ice-free areas or polynyas, brine rejection results from ice formation and dense, saline water sinks to the bottom of the shallow shelf. The sea ice formed in the polynyas is transported southward by the wind. The leading edge melts as it encounters warmer (>0°C) water, thereby cooling and freshening the water column. The thickness, areal coverage, and southernmost extent of sea ice vary on multiple timescales: year-to-year, quasi-decadal, and long-term trends (Niebauer et al., 1999; Wyllie-Echeverria and Ohtani, 1999; Stabeno et al., 2007). In recent decades, the southernmost extent of sea ice was a maximum in the early- to mid-1970s, and a minimum in the early 2000s (Stabeno et al., 2007). In a heavy ice year (e.g., 1975/1976), ice can be advected southward almost to Unimak Pass (54.5°N; Fig. 1) and be present in the north from mid-November until late June. In light ice years (e.g., 2005), the maximum southern extent remains north of the Pribilof Islands (57°N), but can still persist over the northern shelf (north of at 62°N) into June. Recent minima in ice extent are thought to be a response to global climate change and decadal variability (Overland and Stabeno, 2004). The years 2000–2005 were among the warmest on record in the area; winter ocean temperatures stayed above 0°C over the southern shelf (south of 58°N), and in 2005 the depth-averaged water temperature during summer approached 8°C on the southeastern middle shelf. During 2000–2005, shelf temperatures were 2–3°C warmer than those observed in the 1990s and these relatively warm temperatures were a direct response to the atmospheric circulation patterns and reduction in ice cover (Stabeno et al., 2007).
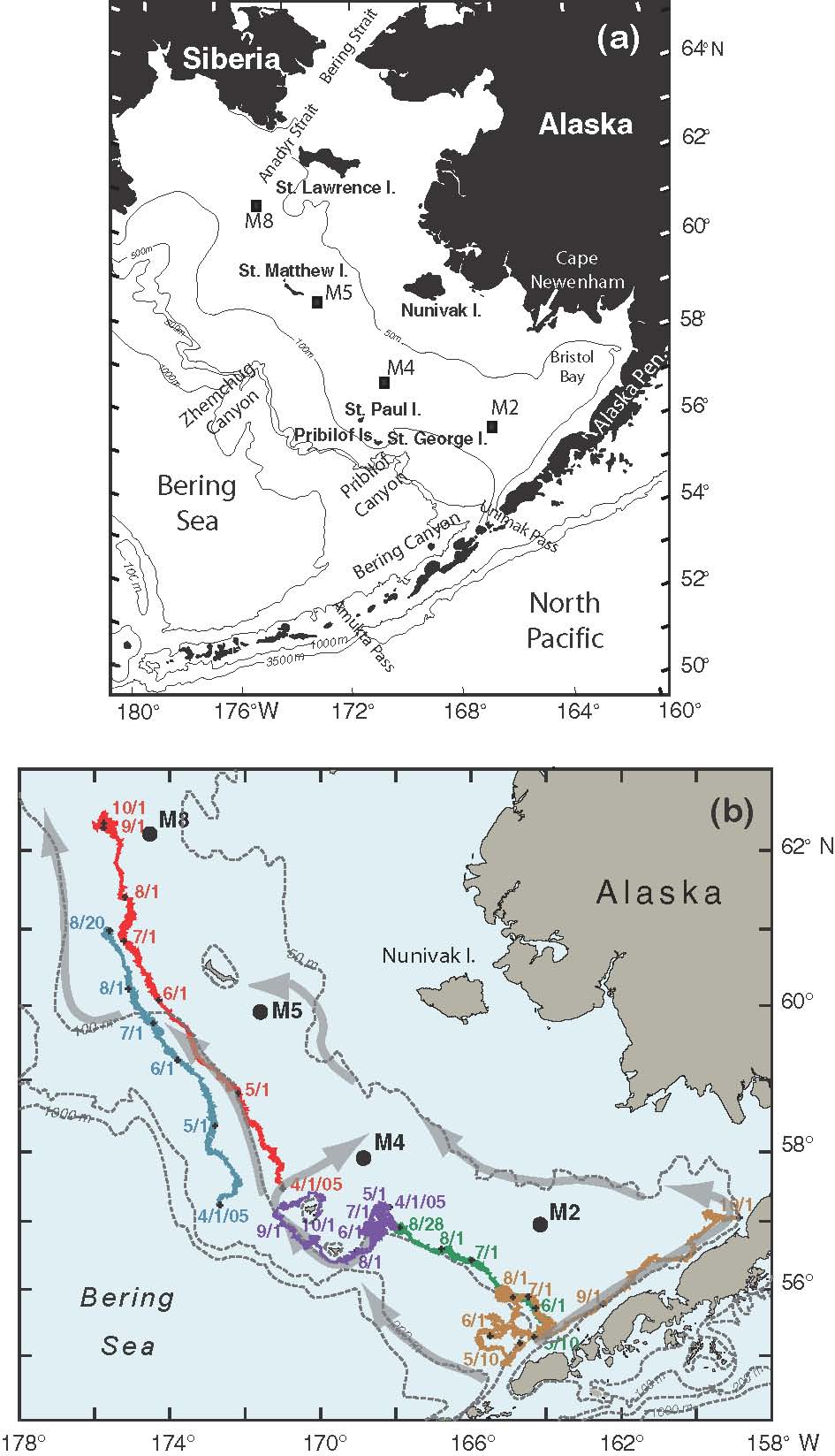
Fig. 1. (a) A map of the study region with bathymetric and geographic labels. (b) The locations of the four primary moorings (M2, M4, M5, and M8) are indicated. The arrows represent the mean flow over the shelf. Also shown are the trajectories from a series of drifters that were deployed in the region. The date of deployment for each drifter is given at the beginning of each trajectory and the position at the beginning of each month is indicated by month/day.
The geographic focus of this paper is the broad eastern shelf. The shelf is ∼500 km wide and slopes gently from the coast westward to the shelf break at ∼180 m. The breadth and flatness of this shelf permits the formation of well-defined, cross-shelf domains. During summer, the eastern shelf south of 62°N is divided into three cross-shelf hydrographic and biological regions: the coastal domain (< 50 m water depth), which is vertically well-mixed by tidal and wind mixing; the middle shelf domain (∼50–100 m), which is sharply stratified into an upper mixed layer (winds) and a lower mixed layer (tides); and the outer shelf domain (∼100–180 m), where the surface wind-mixed layer and the bottom tidally mixed layer are separated by a transitional layer (Coachman, 1986; Kachel et al., 2002). Each of the regions has its own chemical signature (Whitledge et al., 1986; Sullivan et al., 2008), biological communities, and levels of lower trophic-level production (Cooney and Coyle, 1982; Sambrotto et al., 1986; Schneider et al., 1986; Smith and Vidal, 1986; Coyle et al., 1996; Springer et al., 1996).
The eastern Bering Sea shelf also has considerable along-shelf variability, and changes in climate will likely impact the hydrographic and biological differences between the southern and northern regions. For instance, at present, primary production supports a predominantly pelagic food web in the southern portion of the shelf, and a predominantly benthic food web in the northern portion of the shelf (Walsh and McRoy, 1986; Hunt et al., 2002; Grebmeier et al., 2006; Grebmeier and Barry, 2007). Therefore, climate-induced changes in the physical forcing of primary production could have varying effects on these different communities and portions of the shelf depending on the underlying food webs. There are already indications of changes in the northern portion of the shelf south of St. Lawrence Island, and in the Chirikov Basin between St. Lawrence Island and Bering Strait (Grebmeier et al., 2006). Studies begun in the mid-1980s have shown declines in the biomass (Sirenko and Koltun, 1992; Grebmeier, 1993; Grebmeier and Dunton, 2000; Grebmeier et al., 2006) and mean sizes of the dominant bivalves in the area (Lovvorn et al., 2003). The flux of carbon to the benthos in this region is related to a number of factors, including the timing of the ice-edge spring phytoplankton bloom and the seasonal transport of nutrients into the region by the Anadyr Current (Grebmeier et al., 2006).
Our objective for this paper is to describe and compare the underlying mechanisms and processes that distinguish the physics, chemistry, and biology of the northern and southern portions of the eastern middle shelf and the transitional area that lies between them. We consider only that part of the shelf south of ∼62.5°N and exclude the region around and north of St. Lawrence Island which is dominated by the flow through Bering Strait. A latitude of ∼63°N is the northern extent of the middle shelf domain. The area north of that latitude is predominantly shallower than 50 m and the physical and biological processes there may be more similar to the waters in the Chukchi Sea (north of Bering Strait) than they are to processes occurring in the Bering Sea to the south. The ultimate goal is to begin to understand the forces structuring the benthic-dominated northern shelf and the more pelagic-dominated southern shelf so that we can begin to predict how climate change and climate variability will impact the living marine resources of the eastern Bering Sea. Primarily, we use data from two cruises in 2005 (one in the early to mid-spring and the other in late summer/early fall), and data from a series of four moorings on the middle shelf. We begin with an examination of the ice cover and retreat during 2005. Next we compare the spatial patterns in hydrography, chemistry, and biology along the 70-m isobath and how they differed in the spring and late summer. We explore cross-shelf differences using an east–west transect just south of St. Matthew Island. The data from four biophysical moorings provide information on temporal variability of properties between the two cruises. Data from the moorings also reveal the role of currents, especially tidal mixing, on the structure and processes that occur over the shelf.
Go to Next section