The Bering Sea (Fig. 30.1) is a northern extension of the North Pacific Ocean, and is the world's third-largest semi-enclosed sea. A continental shelf underlies about one-half of the Bering Sea, consisting of a broad (>500 km) eastern portion between the Alaska Peninsula and Cape Navarin, and a narrower (<100 km) western portion between Cape Navarin and Kamchatka Strait. Mean northward transport (~0.8 × 10![]() m
m![]() s
s![]() ) through Bering Strait is driven by an ~0.5 m sea level difference between the Bering Sea and Arctic Ocean (Coachman, 1993). This transport provides the only connection and exchange of water between the Pacific and Atlantic Oceans in the northern hemisphere. This northward flow requires transport onto the shelf of nutrient-rich slope waters from the Bering Slope Current, the eastern boundary current of the cyclonic gyre that dominates circulation in the Bering Sea gyre (Stabeno et al., 1999). Over the northern shelf, nutrient-rich water upwells due to shoaling topography, thereby stimulating primary production (Nihoul et al., 1993). Ice forms predominantly in the polynyas over the northern shelf. The cold, saline water formed from brine rejection sinks and flows northward through Bering Strait. Globally, this water plays a role in maintaining the Arctic Ocean halocline and hence ventilation of the deep waters (Aagaard et al., 1985).
) through Bering Strait is driven by an ~0.5 m sea level difference between the Bering Sea and Arctic Ocean (Coachman, 1993). This transport provides the only connection and exchange of water between the Pacific and Atlantic Oceans in the northern hemisphere. This northward flow requires transport onto the shelf of nutrient-rich slope waters from the Bering Slope Current, the eastern boundary current of the cyclonic gyre that dominates circulation in the Bering Sea gyre (Stabeno et al., 1999). Over the northern shelf, nutrient-rich water upwells due to shoaling topography, thereby stimulating primary production (Nihoul et al., 1993). Ice forms predominantly in the polynyas over the northern shelf. The cold, saline water formed from brine rejection sinks and flows northward through Bering Strait. Globally, this water plays a role in maintaining the Arctic Ocean halocline and hence ventilation of the deep waters (Aagaard et al., 1985).
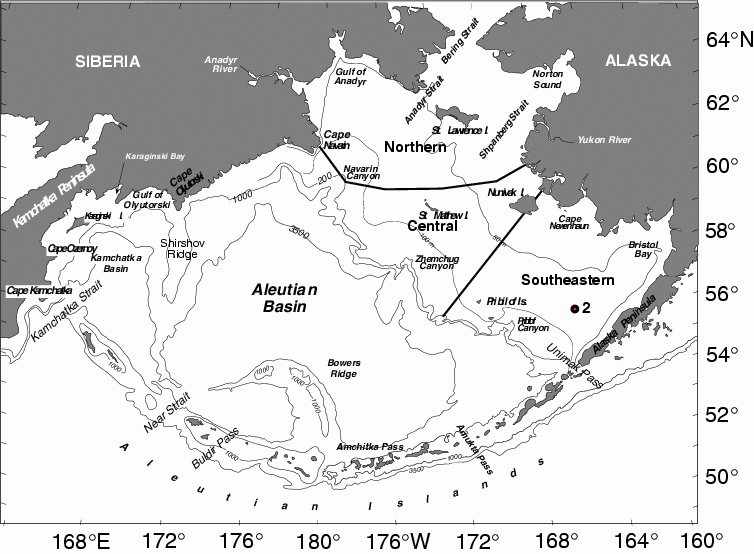
Figure 30.1: Geography and place names of the Bering Sea. The location of the M2 is indicated. Depth contours are in meters. (From Schumacher and Stabeno, 1998 or Stabeno et al., 1999.)
The eastern Bering Sea is home to a rich variety of biota, including the world's most extensive eelgrass beds; at least 450 species of fish, crustaceans, and mollusks; 50 species of seabirds; and 25 species of marine mammals. It has been classified as its own bio-geographic province (Longhurst, 1998) and as a Class II, moderately high (150–300 gC/m![]() /yr) productivity, ecosystem based on satellite remote sensing estimates (www.edc.uri.edu/lme/text/east-bering-sea.htm), although primary production estimates for sub-regions are much higher (e.g., Springer and McRoy, 1993). The abundant fish and wildlife of the Bering Sea have supported the lives and livelihoods of Asians and North Americans since prehistoric times. For the 65,000 Alaskan natives living on the shores of the eastern Bering Sea (NRC, 1996), subsistence is not just a means of providing food; it is the wellspring of spiritual and cultural traditions. Presently (2002), the U.S. Bering Sea fishery provides about 40% of the U.S. and about 5% of the world harvest of fish and shellfish with walleye pollock (Theragra chalcogramma) comprising much of the fish landings; the tanner crab (Chionoecetes bairdi) fishery currently is the largest crustacean fishery (by weight) in the U.S. Bristol Bay and supports the world's largest sockeye salmon (Onchorhynchus nerka) fishery. In addition to supporting a large portion of the nation's fishery production, the Bering Sea supports ~80% of the U.S. nesting seabird population, comprising approximately 36 million birds. Furthermore, many endemic species such as red-legged kittiwakes (Rissa brevirostris) and whiskered auklets (Aethia pygmaea) are found in the Bering Sea and further highlight the significance of this region. Prior to the 1960s, there were much larger populations of northern fur seals (Callorhinus ursinus), Steller sea lions (Eumetopias jubatas), whales and birds. The U.S. government has listed the Steller sea lion and spectacled eider (Somateria fischeri) as endangered species, sea otters (Enhydra lutris) as threatened, and there are declines of some salmon populations that have adversely affected commercial and subsistence harvest. This region's wetlands, coastlines and islands provide globally significant habitats for many additional wildlife species, and its natural history holds answers to important questions about world history, including the introduction of hominids to North America.
/yr) productivity, ecosystem based on satellite remote sensing estimates (www.edc.uri.edu/lme/text/east-bering-sea.htm), although primary production estimates for sub-regions are much higher (e.g., Springer and McRoy, 1993). The abundant fish and wildlife of the Bering Sea have supported the lives and livelihoods of Asians and North Americans since prehistoric times. For the 65,000 Alaskan natives living on the shores of the eastern Bering Sea (NRC, 1996), subsistence is not just a means of providing food; it is the wellspring of spiritual and cultural traditions. Presently (2002), the U.S. Bering Sea fishery provides about 40% of the U.S. and about 5% of the world harvest of fish and shellfish with walleye pollock (Theragra chalcogramma) comprising much of the fish landings; the tanner crab (Chionoecetes bairdi) fishery currently is the largest crustacean fishery (by weight) in the U.S. Bristol Bay and supports the world's largest sockeye salmon (Onchorhynchus nerka) fishery. In addition to supporting a large portion of the nation's fishery production, the Bering Sea supports ~80% of the U.S. nesting seabird population, comprising approximately 36 million birds. Furthermore, many endemic species such as red-legged kittiwakes (Rissa brevirostris) and whiskered auklets (Aethia pygmaea) are found in the Bering Sea and further highlight the significance of this region. Prior to the 1960s, there were much larger populations of northern fur seals (Callorhinus ursinus), Steller sea lions (Eumetopias jubatas), whales and birds. The U.S. government has listed the Steller sea lion and spectacled eider (Somateria fischeri) as endangered species, sea otters (Enhydra lutris) as threatened, and there are declines of some salmon populations that have adversely affected commercial and subsistence harvest. This region's wetlands, coastlines and islands provide globally significant habitats for many additional wildlife species, and its natural history holds answers to important questions about world history, including the introduction of hominids to North America.
The current state of this ecosystem is a product of its history that includes substantial human and natural impacts. Western knowledge of the Bering Sea's rich ecosystem began with Vitus Bering's voyage in 1741 (Ford, 1966). Over the next two centuries, commercial exploitation brought the Steller sea cow to extinction, and the sea otter, fur seal, walrus and bowhead whale nearly to extinction (Fay, 1981). In the early 1950s, the "commercial fishing period" began and that, together with the removal of marine mammals (until the early 1970s), had a marked impact on the composition of the ecosystem (NRC, 1996). These factors form the basis of the Trophic Cascade Hypothesis (Merrick, 1997; NRC, 1996). As the large populations of mammals (particularly whales) and fish were removed, there was an increased amount of food (zooplankton and small fish) for other vertebrate predators. There is little doubt that the over-exploitation and reductions of these populations, together with a regime shift in the physical environment, led to a switch in the late 1960s and early 1970s to a system dominated by pollock, which has persisted since then (NRC, 1996).
Much of the early oceanographic research was conducted to address questions relating to the international fishery (Takenouti and Ohtani, 1974; Arsenev, 1967; Hood and Kelley, 1974; and Favorite et al., 1976). In the early 1950s, Hokkaido University began annual training cruises to the Bering Sea and established what has become one of the longest physical and biological time series available in the region (e.g., Sugimoto and Tadokoro, 1997). Between the mid-1970s and late 1980s, the focus changed to resource assessment on the eastern shelf as part of the Outer Continental Shelf Environmental Assessment Program (OCSEAP; Hood and Calder, 1981). Annual groundfish and crab assessment continues to this day by NOAA/National Marine Fisheries Service. OCSEAP was followed by ecosystem research over the southeastern (PROBES; Processes and Resources of the Bering Sea: Hood, 1999) and northern shelf (ISHTAR; Inner Shelf Transfer and Recycling in the Bering and Chukchi Seas: McRoy, 1999), and examination of ice related phenomenon during the Bering Sea Marginal Ice Zone Experiment (MIZEX: Muench, 1983). During the 1990s, research focused again on fisheries and the influence of the physical factors on the ecosystem. Major contributions included those from the Fisheries Oceanography Coordinated Investigations program (FOCI: Schumacher and Kendall, 1995); NOAA's Coastal Ocean Programs, Bering Sea FOCI (BSFOCI: Bailey et al., 1999; Brodeur et al., 1999b; Napp et al., 2000) and the Southeast Bering Sea Carrying Capacity program (SEBSCC: Stabeno and Hunt, 2002; Macklin et al., 2002); the National Science Foundation (Polar Programs) "Study of prolonged production, trophic transfer, and processes at the Bering Sea inner front" (hereafter called Inner Front Study: Stabeno and Hunt, 2002) and a program to conduct long-term ecological research on marine ecosystems in the Arctic and Pacific Oceans (BERPAC: Tsyban, 1999). Alexander (1999) provides a review of many of the interdisciplinary studies of the Bering Sea.
Features of the Bering Sea (including solar radiation, atmospheric phenomenon, sea ice cover, and water column structure and temperature, and biological production) vary over a wide range of time scales. These include strong seasonal and annual signals and smaller, but potentially important longer-period fluctuations. Over the vast region of the eastern shelf, most of the physical phenomena also vary greatly with both latitude and longitude. The variability in physical features, together with the flux of nutrient-rich slope waters onto the eastern shelf, shapes one of the world's most productive ecosystems (Walsh et al., 1989). Observations suggest that a highly productive region also exists over the edge of the continental shelf (Springer et al., 1996). This "green belt" extends around the entire perimeter of the continental shelf and estimates of both primary and secondary production in this belt are more than 60% greater than production over the adjacent shelf waters.
In this chapter, we first present a conceptual model of the pathways by which changes in atmospheric phenomenon can influence sea ice, oceanic features and biota. While some of the pathways may be specific to the Bering Sea, much is directly applicable to other subarctic seas. We then present the salient features of the physical environment summarized from Schumacher and Stabeno (1998), updated to include more recent findings (e.g., Kachel et al., 2002; Overland et al., 1999a; Stabeno et al., 2002). Our focus here is on interdisciplinary ocean science, with a particular focus on several phenomena whose influence on biota has been established: water temperature, transport and turbulence. Following Schumacher and Stabeno (1998), we partition the continental shelf of the Bering Sea into eastern (primarily U.S. waters) and western regions, with the former further differentiated into southeastern, central and northern sub-regions. The following sections describe processes of the eastern Bering Sea shelf and those over the northern and western shelf. We conclude with a discussion of suggestions for future research.
The complex pathways that weave together the biotic and abiotic components of the Bering Sea ecosystem are not all known, nor are all the interactions among the various components well understood. Yet, identifying and understanding mechanisms that transfer climate change in the atmosphere and ocean to biota is essential if we are to comprehend ecosystem dynamics (Francis et al., 1998). Fluctuations in the physical environment can impact the ecosystem through both changes in the nutrient-phytoplankton-zooplankton sequence (i.e., bottom up control), and/or by altering habitat or some other condition that results in changes in abundance and/or spatial composition of higher trophic level animals (i.e., top down control). Temperature, turbulence and transport are mechanisms through which the physics influences the biology. These, in turn, respond to wind and, for high latitude seas, sea ice. All of these phenomena are included in a conceptual model (Fig. 30.2) that provides a schematic of the pathways in which changes in the physical environment impact biota. While the sequence is generally thought of as unidirectional starting with changes in the atmospheric climate driving those in the ocean and on to biota, changes in oceanic environment (e.g., heat fluxes) can alter atmospheric features (a feedback loop). In this conceptual model, the direct influence of the physical environment is a function of trophic level; the arrows between levels represent both biological interactions and the communication of physical influences.
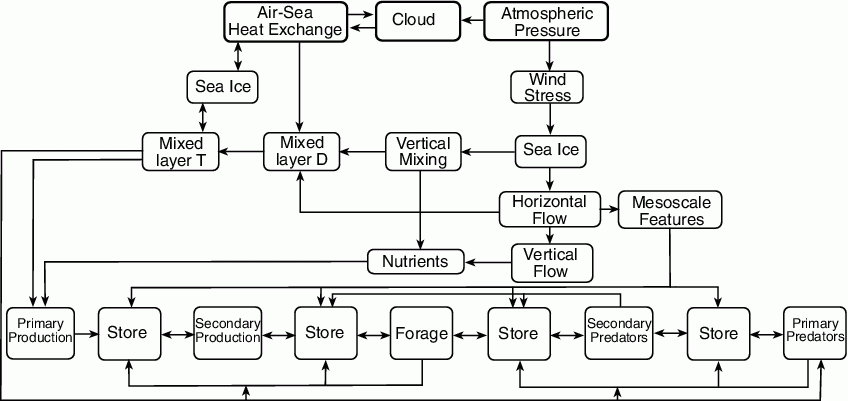
Figure 30.2: Pathways whereby changes in physical environment influence the biological processes (after Schumacher et al., 2003). Sea ice appears twice because it represents pathways for different processes.
The pathways in the conceptual model also include biological interactions (e.g., density dependence) that can be a dominant factor in population dynamics; for example, cannibalism has a marked influence on recruitment processes of pollock in the eastern Bering Sea (Livingston and Methot, 1998). In a recent examination of variations in the strength of pollock year classes (Wespestad et al., 2000), physical and biological mechanisms set survival. For instance, the availability of young pollock as prey for adult fish may be related to transport of larval stage animals. A switch model of larval pollock dynamics (e.g., Napp et al., 2000) also highlights that physical and biological factors interact to influence survival.
Understanding the influence of changes in the physical environment on the ecosystem is confounded by many factors. Physical conditions favorable for one life history stage may be detrimental for another. In addition, many marine populations are likely to respond to change in a nonlinear fashion (e.g., Bailey et al., 2005; Cury et al., 1995; Stenseth et al., 2002). For example, it is hypothesized that during cold years in the Bering Sea, the spatial domain of age-1 pollock over the shelf is restricted to warmer waters (a behavioral response to temperature) and that increased local densities of these fish result in higher predation mortality by older fish (Ohtani and Azumaya, 1995). Cold winters with extensive ice cover can also provide conditions that support rapid increases of primary production associated with the presence of sea ice. Due to the cold-water temperatures and related low physiological rates, the limited zooplankton biomass is unable to consume a significant portion of the primary production. Thus, most of the carbon sinks to the sea floor, feeding the benthic food web (Walsh and McRoy, 1986). In addition, it has been demonstrated that nitrate uptake on the southeastern Bering Sea shelf has a nonlinear relationship to wind induced mixing, and timing of storms relative to the phase of the production system (i.e., respiration or nutrient limited period) is critical (Sambrotto et al., 1986).
While many environmental features have been well documented (e.g., Schumacher and Stabeno, 1998; Stabeno et al., 2001; Hunt et al., 2002), temporal and spatial variations in the amount of solar radiation and its potential impact on ecosystem dynamics has not been well addressed, yet such variations are evident in time series of net short wave radiation (Fig. 30.3). The effect of latitude is apparent in these time series, with the northernmost station receiving approximately 19% less energy. The other two net short-wave radiation time series, which are at similar latitudes, appear coherent. The northern and the two southern time series show similar patterns of variability in 1972–1983 and 1995–2001. During the intervening period the time series appear to be out of phase by ~180 degrees with the magnitude of variability of the northern series much less than that seen in the southern time series. While the two series at approximately the same latitude are often similar in magnitude, there are some years (e.g., 1995) when they differ by more than 10 Wm![]() . Some of the longer-term variation may be due to the sun spot cycle that results in a solar flux variation of ~0.1% or ~2 Wm
. Some of the longer-term variation may be due to the sun spot cycle that results in a solar flux variation of ~0.1% or ~2 Wm![]() at sea level. It is likely, however, that regional differences in cloud cover account for the majority of this signal. It is not presently known how these differences influence ecosystem dynamics, although the latitudinal differences relate to a shorter more intense period of production over the northern shelf than exists over the southeastern shelf. This likely contributes to the observed differences in the dominant pathway of carbon cycling (pelagic versus benthic): the northern shelf is predominately a benthic system, whereas the southeastern shelf may be either pelagic or benthic depending on timing of spring blooms (Walsh and McRoy, 1986; McRoy, 1993).
at sea level. It is likely, however, that regional differences in cloud cover account for the majority of this signal. It is not presently known how these differences influence ecosystem dynamics, although the latitudinal differences relate to a shorter more intense period of production over the northern shelf than exists over the southeastern shelf. This likely contributes to the observed differences in the dominant pathway of carbon cycling (pelagic versus benthic): the northern shelf is predominately a benthic system, whereas the southeastern shelf may be either pelagic or benthic depending on timing of spring blooms (Walsh and McRoy, 1986; McRoy, 1993).
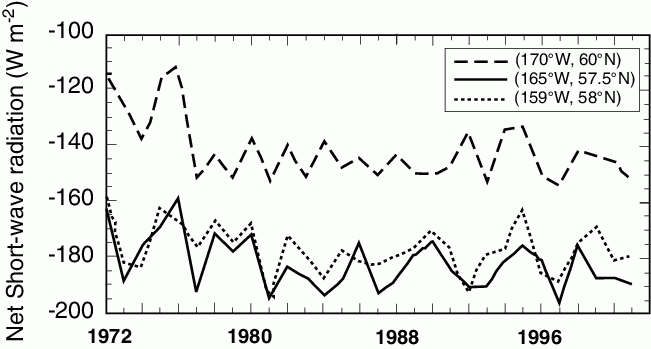
Figure 30.3: Average net short wave radiation (NSWR) from April 1–June 30 (Wm![]() ). Note negative indicates a downward flux.
). Note negative indicates a downward flux.
Recent research (e.g., Minobe, 1999, 2002; Hare and Mantua, 2000) has highlighted atmospheric variations that exist on multi-decadal time scales (typically, 10 to 70 years), which cause significant alterations throughout the ecosystem. These changes, known as regime shifts, are seen in the biological environment primarily as changes in community composition and/or biomass of a given species. The mechanisms that initiate a climate regime shift, however, are presently unknown. In a review of how climate variability impacts biota in the eastern Bering Sea, Schumacher and Alexander (1999) identify the following four candidates for forcing interdecadal signals in that region. First, changes in solar activity are correlated with temperature and pressure in the atmosphere over the North Pole (Labitzke and van Loon, 1988), and may provide a forcing mechanism for decadal oscillations in the coupled air-ice-sea system in the northern hemisphere (Ikeda, 1990). The lunar nodal cycle of the moon (period of 18.6 yrs) affects mixed layer depths and has been linked to oceanic conditions in the North Pacific Ocean (Royer, 1998; Royer et al., 2001). Third, atmospheric interactions exist between the Southern Oscillation and the Aleutian Low as proposed by Wooster and Hollowed (1995). Finally, Latif and Barnett (1994) suggest that an unstable air-sea interaction exists between circulation in the North Pacific subtropical gyre and the Aleutian Low pressure system. Since 1999, other candidates have been proposed, including changes in mid-latitude wind stress (Parrish et al., 2000), subtropical oceanic Rossby waves (Jin et al., 2001), and subsurface changes in the ocean south of the equator as part of an oscillation intrinsic to the equatorial and southern tropical Pacific Ocean (Giese et al., 2002).
The primary atmospheric pressure features influencing the Bering Sea include weather patterns in the tropical South Pacific (El Niño-Southern Oscillation: ENSO), North Pacific (Pacific-North America: PNA) and Arctic (Arctic Oscillation: AO). The mode of connectivity between these hemispheric-scale features and the regional weather appears to be mainly a perturbation in the magnitude, pathway, and frequency of storm passage along the Aleutian Island chain (Stabeno et al., 1999). The frequent migration of storms results in a statistical feature known as the Aleutian Low. During summer with its long periods of daylight and high solar radiation, the Aleutian Low is typically weak and the weather benign. During winter, a marked change occurs in atmospheric pressure fields. High sea level pressure (Siberian High) dominates Asia, while the Aleutian Low deepens and dominates weather over the North Pacific and Bering Sea. The juxtaposition of these features results in strong, frigid winds from the northeast. The frequency and intensity of storms in the southern Bering Sea decreases temporally from winter to summer, but the frequency also decreases with increasing latitude (Overland, 1981; Overland and Pease, 1982).
The physical features of the oceanic and shelf region of the eastern Bering Sea were reviewed by Schumacher and Stabeno (1998) and Stabeno et al. (1999). Over the southeastern shelf during summer, three distinct cross shelf domains exist which are characterized by water column structure, currents and biota (Cooney and Coyle, 1982; Coachman, 1986; Schumacher and Stabeno, 1998). These are the coastal (depth <50 m and characterized by weak stratification), middle shelf (50–100 m deep, characterized by a wind-mixed surface layer abutting a tidally mixed bottom layer), and outer shelf (100–180 m deep, mixed upper and lower layers separated by a layer with slowly increasing density). Over much of the southeastern shelf during summer, temperature is an excellent indicator of vertical water-column structure. The primary source of freshwater is the melting of sea ice; the sea ice is advected southward during the winter and early spring. During early spring, the combination of tidal and wind mixing tend to weaken any strong halocline. For waters <30 m deep, tidal mixing energy tends to stir the entire water column. In the deeper waters (30 < z < 90 m), there is some vertical structure in salinity, but it is typically <0.3 psu (Overland et al., 1999b). It is only in late spring and summer when the storms weaken and solar heating begins, that pronounced vertical stratification occurs over the middle domain. During winter, the domains are poorly defined since storms mix the water column to >90 m.
These summer (April-October) domains are separated by a system of transitional zones or fronts (Coachman, 1986; Iverson et al., 1979; Schumacher and Stabeno, 1998). The shelf-break front separates the outer shelf from slope waters; the broad middle shelf transition zone lies between outer and middle shelf waters; and an inner (structure) front separates the well-mixed coastal waters and the two-layered middle shelf domain. Knowledge of the characteristics of the inner front was recently refined: the inner front is wider than previously thought and its location varies by tens of kilometers (Kachel et al., 2002), rather than being relatively fixed to the 50-m isobath as earlier hypothesized. The balance of wind and tidal energy plays a major role in shaping the vertical structure of both the coastal and middle shelf domains (Schumacher and Stabeno, 1998; Coachman, 1986). The domains provide unique habitats for biota. For example, the meso-zooplankton community in the two shallower domains is comprised primarily of the small to medium-sized copepods, whereas in the outer shelf domain and oceanic region, large copepods dominate (Cooney and Coyle, 1982; Smith and Vidal, 1986).
We consider the northern shelf to be that portion of the eastern shelf north of ~62°N where changes in topography, tidal energy, and river discharge (primarily from the Yukon River) modify the boundaries between domains. The width of both the coastal and middle shelf domains increase. Nearly the entire shelf east of the Anadyr Strait has depths <50 m, and much of the Gulf of Anadyr lies in the middle domain. North of Nunivak Island, the inner front moves to the vicinity of the 30-m isobath as tidal mixing energy decreases (Schumacher and Stabeno, 1998). In the vicinity of the Yukon River delta, however, the substantial freshwater discharge can result in stratification in waters <30 m. The water column (generally <20 m) in Norton Sound typically exhibits a two-layered structure during summer as opposed to the coastal domain in the southeast, which is at most weakly stratified. During winter, strong heat and salt fluxes result in a vertically mixed water column (Muench et al., 1981).
Across the shelf south of St. Lawrence Island, three water masses exist: Alaskan Coastal, Bering Shelf, and Anadyr (Coachman et al., 1975). The accompanying regional salinity field is characterized by a zonal gradient with salinity decreasing from east to west (Coachman et al., 1975; Schumacher et al., 1983). In water where the temperature is <0°C, the salinity between ~30 to 60 m increases by 2.5 psu from east to west across the shelf. During ice-free conditions, salinity from St. Lawrence Island to the Gulf of Anadyr seldom exceeds 32.8 psu; however, during ice formation values >34.2 psu occur. In the Gulf of Anadyr, salinity during summer ranges between 33.0 and 33.5 psu. The saline waters which flow northwestward across the mouth of the Gulf of Anadyr carry relatively warm temperatures and nutrients which suggests the presence of outer shelf water. North of St. Lawrence Island, all three water masses are present and can be identified as they flow northward through Bering Strait (Coachman et al., 1975).
A system of three hydrological zones exists over the western shelf that are somewhat analogous to those found on the eastern shelf (Verkhunov, 1994). These hydrological zones (coastal, transitional and oceanic) are easily distinguished in temperature/salinity diagrams (Khen, 1999). The coastal zone has low (<31.0 psu) salinity surface water and a strong pycnocline. Due to a less pronounced halocline, the transitional zone exhibits a more weakly two-layered structure than the coastal zone. In both zones, a strong seasonal thermocline develops over summer. The oceanic zone is identified by a three-layered vertical structure with relatively warm (~1.0°C) bottom temperatures that indicate the presence of slope waters.
While there is some similarity to the hydrographic structure of the eastern shelf, fundamental differences exist. The locations of the zones over the western shelf are not stationary, although they can at times be associated with depth contours as occurs on the eastern shelf (Verkhunov, 1994). The western shelf is relatively narrow and divided by peninsulas into three separate and somewhat isolated gulfs. The immense width of the eastern shelf eliminates the direct influence of gyre circulation on all but the outer domain. Such is not the case on the western side, where the Kamchatka Current, the western boundary current of the Bering Sea gyre, has a profound impact on the location of hydrographic zones. When this current moves shoreward and flows over the continental slope, all the zones are compressed and their frontal divisions are shifted shoreward (Khen, 1999).
A schematic of the general circulation is shown in Fig. 30.4. The portion of the Alaskan Stream that flows through Aleutian passes, especially Amchitka and Amukta Passes, forms the eastward flowing Aleutian North Slope Current (ANSC: Reed and Stabeno, 1999; Stabeno et al., 1999). The ANSC provides the main source of the Bering Slope Current (BSC), which is often characterized as variable flow replete with eddies and meanders, although at times it appears as a more organized northwestward flowing current (Stabeno et al., 1999). The BSC separates from the slope, resulting in a broad weak westward flow across the basin. Along the west coast the current intensifies, forming the southward flowing Kamchatka Current. Exchange between the slope and eastern shelf likely varies depending upon which mode of the BSC is dominant. Transport in the ANSC/BSC system is highly variable ranging from ~2 × 10![]() m
m![]() s
s![]() to ~9 × 10
to ~9 × 10![]() m
m![]() s
s![]() (Stabeno et al., 1999). The importance of these slope currents to ecosystem dynamics of the eastern shelf is threefold. First, they transport material northward along the slope that can be advected onto the shelf through various onshelf mechanisms (Schumacher and Stabeno, 1994; Stabeno et al., 2001). Second, eddies, which are common seaward of the shelf break (Schumacher and Reed, 1992), apparently provide a temporary habitat that favors survival of larvae (Schumacher and Stabeno, 1994) and are regions of enhanced primary production (Mizobata et al., 2002). Finally, the vertical temperature structure of the slope current with its warm subsurface maximum is potentially important to fish stocks (Reed, 1995).
(Stabeno et al., 1999). The importance of these slope currents to ecosystem dynamics of the eastern shelf is threefold. First, they transport material northward along the slope that can be advected onto the shelf through various onshelf mechanisms (Schumacher and Stabeno, 1994; Stabeno et al., 2001). Second, eddies, which are common seaward of the shelf break (Schumacher and Reed, 1992), apparently provide a temporary habitat that favors survival of larvae (Schumacher and Stabeno, 1994) and are regions of enhanced primary production (Mizobata et al., 2002). Finally, the vertical temperature structure of the slope current with its warm subsurface maximum is potentially important to fish stocks (Reed, 1995).
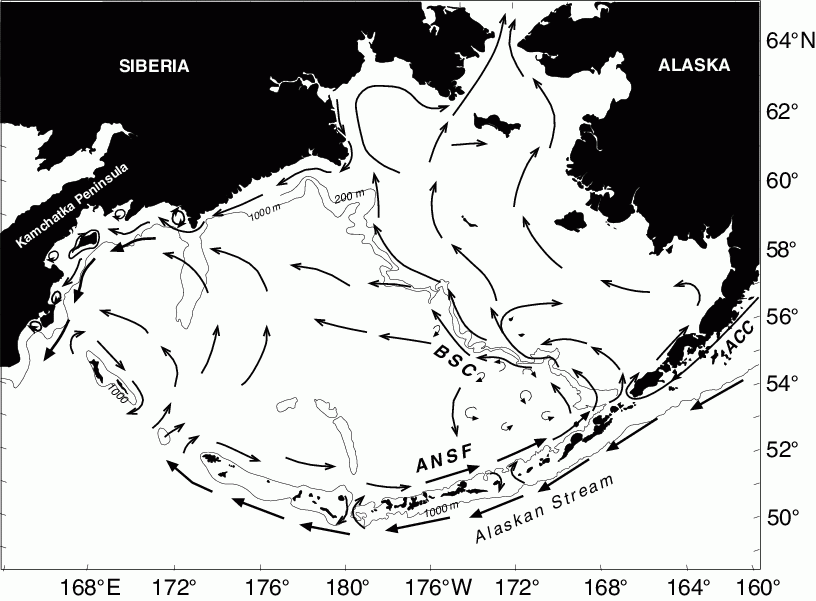
Figure 30.4: Schematic of mean circulation in the upper 40 m over the basin and shelf (from Stabeno et al., 1999). The arrows with large heads represent currents with mean speeds >50 cm s![]() . The Alaskan Stream, Aleutian North Slope Current (ANSC), Bering Slope Current (BSC) and Kamchatka Current are indicated. Depth contours indicate 1000 m isobath and in the Bering Sea the 200 m isobath.
. The Alaskan Stream, Aleutian North Slope Current (ANSC), Bering Slope Current (BSC) and Kamchatka Current are indicated. Depth contours indicate 1000 m isobath and in the Bering Sea the 200 m isobath.
Low frequency flow on the shelves is much weaker than in the basin. Over the eastern shelf, the net flow from late spring through mid autumn is northward. Enhanced, more organized flow occurs along the 100 m isobath and the 50 m isobath. Both of these coincide with frontal or transition zones on the shelf. The flow along the 50 m isobath, which we call the Bering Coastal Current (BCC), is weak (1–5 cm s![]() ). The BCC consists of waters from the Gulf of Alaska's Alaska Coastal Current (ACC) that flows through Unimak Pass with a transport of ~0.3 × 10
). The BCC consists of waters from the Gulf of Alaska's Alaska Coastal Current (ACC) that flows through Unimak Pass with a transport of ~0.3 × 10![]() m
m![]() s
s![]() (Schumacher and Stabeno, 1998; Stabeno et al., 2002). Some of this transport flows along the Alaska Peninsula and the west coast of Alaska where freshwater is added from widely distributed riverine sources. Unlike other buoyancy driven flows that are strengthened and compressed to the coastline by alongshore wind stress (e.g., the ACC; Stabeno et al., 1995), the BCC is strongest in the vicinity of the 50 m isobath, which can be more than 100 km offshore (Schumacher and Stabeno, 1998; Schumacher and Kinder, 1983). After flowing around the entire perimeter of the eastern shelf, the BCC exits the Bering Sea through Bering Strait.
(Schumacher and Stabeno, 1998; Stabeno et al., 2002). Some of this transport flows along the Alaska Peninsula and the west coast of Alaska where freshwater is added from widely distributed riverine sources. Unlike other buoyancy driven flows that are strengthened and compressed to the coastline by alongshore wind stress (e.g., the ACC; Stabeno et al., 1995), the BCC is strongest in the vicinity of the 50 m isobath, which can be more than 100 km offshore (Schumacher and Stabeno, 1998; Schumacher and Kinder, 1983). After flowing around the entire perimeter of the eastern shelf, the BCC exits the Bering Sea through Bering Strait.
While some of the transport through Unimak Pass forms the BCC, the remainder flows northwestward in the vicinity of the 100 m isobath to the Pribilof Islands. The strongest mean current over the central and southeastern shelf is found south of St. George Island, where the outer shelf narrows and the bottom slope increases and thus narrowing the flow along the 100 m isobath. Daily average currents can exceed 40 cm s![]() . The mean flow over the rest of the central and southeastern shelf is weak (generally <5 cm s
. The mean flow over the rest of the central and southeastern shelf is weak (generally <5 cm s![]() ). The currents over the northern shelf are stronger, especially the Anadyr Current which is the primary source of water flowing through Bering Strait. During winter, the flow becomes less organized over the shelf as the frontal structures break down and strong winter storms mix the water to the bottom.
). The currents over the northern shelf are stronger, especially the Anadyr Current which is the primary source of water flowing through Bering Strait. During winter, the flow becomes less organized over the shelf as the frontal structures break down and strong winter storms mix the water to the bottom.
Tidal currents are a major circulation feature of the southeastern shelf, but over the northern and western shelves their importance is greatly diminished (Schumacher and Stabeno, 1998; Stabeno et al., 1999; Kowalik and Stabeno, 1999). Over the southeastern shelf, tides are the dominant source of kinetic energy and they typically mix the coastal waters (z < ~50 m) and the lower 30–40 m of the deeper portions of the shelf. In addition, their interaction with bathymetric features can result in residual flow, particularly in canyons (Schumacher and Reed, 1992; Kowalik and Stabeno, 1999) and around islands such as the Pribilof Islands (Kowalik and Stabeno, 1999; Stabeno et al., 1999).
The eastern Bering shelf is a marginal ice zone, and sea ice markedly impacts the ecosystem. Ice-free conditions typically exist from June through October. Formation of sea ice generally begins in the polynyas of the northern shelf in November, with maximum ice extent occurring as early as January or as late as April, but more typically in March. The mechanism of ice formation for the eastern Bering Sea has been described by analogy to a "conveyor belt" (Overland and Pease, 1982). Ice is produced along leeward (south-facing) coasts of the northern shelf, and is driven southward by wind to the vicinity of its thermodynamic limit where it melts. This limit advances southward as cold northerly winds and ice-melt cool the shelf waters. The amount of production and advection of ice depends upon which storm track dominates, with greatest ice production occurring in years when the Aleutian Low is well developed and displaced eastward so that storms migrate along a primary storm track south of the Alaska Peninsula. Satellite observations of ice cover show ~40% variation about the mean (Niebauer, 1998). Using the same sea ice data set, but resolved along a longitude, Wyllie-Echeverria (1995) showed that other characteristics also exhibit large interannual variability. These characteristics include: duration of ice at its southern extent (3–15 weeks), time of retreat from the southernmost extent (between February and June), and the number of weeks that ice remained over the middle shelf (3–28 weeks, with a mean of 20).
Changes in characteristics of sea ice over the southeastern shelf (including the time of arrival, departure and persistence), indicate that the most extensive ice years coincided with the negative phase of the Pacific Decadal Oscillation (the first mode of decadal variability in sea surface temperature over the North Pacific Ocean), although considerable interannual variability occurs (Stabeno et al., 1998 and 2001). These authors characterized temporal variability in spatial patterns by dividing the time series of ice observations into three subsets according to generally agreed upon periods: 1972–76 (cold), 1977–88 (warm), and 1989–98 (weaker cold). A marked difference exists in persistence and spatial distribution of ice between the first and the latter two periods. During the cold period, ice covered the shelf out to and over the slope, and remained around St. Paul Island for more than a month. During the later years, ice did not extend as far seaward and its residence time was typically 2–4 weeks less than during the cold period. The differences between the two latter regimes are more subtle, but still evident. Between the mid-shelf and slope northwest of the Pribilof Islands, ice remained for 2–4 weeks longer during 1989–1998 than during 1977–1988. North and west of St. Lawrence Island and along the coast of Alaska north of Kuskokwim Bay, there were 1–2 less weeks of ice cover in 1989–1998 than in 1977–1988. Changes in the Arctic Oscillation and the attendant changes in wind patterns are likely the causal mechanism for these recent alterations in sea ice (Stabeno and Overland, 2001). In addition, marked differences existed in the ice distribution and extent along the Alaskan Peninsula. During the cold period, ice extended seaward nearly to Unimak Pass, whereas in the other periods contours of ice persistence were constrained or limited to inner Bristol Bay. This pattern is likely related to variations of inflow and/or temperature of shelf waters from the Gulf of Alaska, which flows through Unimak Pass onto the Bering Sea shelf (Schumacher and Stabeno, 1998).
Cold northerly winds, which advect the ice southward, and melting play a critical role in fluxes of heat and salt, and in generation of both baroclinic flow and the cold lower layer (cold pool) which persists all summer over the middle shelf domain (Schumacher and Stabeno, 1998; Wyllie-Echeverria and Ohtani, 1999). The positive buoyancy from melting ice initiates both baroclinic transport along the marginal ice zone and stratification. Cooling and mixing associated with ice-advance help to condition the entire water column over the middle shelf domain (Stabeno et al., 1998). With subsequent seasonal heating of the upper layer, the lower layer becomes insulated and temperatures often remain below 2°C (Reed, 1995). The annual area of this cold pool varies by ~2.0 × 10![]() km
km![]() between maximum and minimum extent.
between maximum and minimum extent.
Natural fluctuations in climate dramatically influence biota in the Bering Sea (e.g., Napp and Hunt, 2001). Trends observed in 11 species (fish, marine birds, and marine mammals) found in the eastern Bering Sea/Aleutian Islands ecosystem show marked changes in relative abundance (Fig. 30.5). Using 100 physical and biological time series (29 of these from eastern Bering Sea marine biota), Hare and Mantua (2000) showed a significant change occurred in 1976/77 and to a lesser degree in 1989. While the mechanisms that link climate to biota were not addressed, it appears that a shift in the decadal patterns of climate (indicated by changes in the Pacific Decadal Oscillation and the Arctic Oscillation) contributed to changes in biota. More recently the change in the Pacific Decadal Oscillation (PDO) in the late 1990s has been interpreted as a regime shift (McFarlane et al., 2000; Macklin et al., 2002; Peterson and Schwing, 2003) or more recently as a shift in the dominate mode of variability in sea surface temperature (Bond et al., 2004). It is not necessary for the climate shift to be immediately manifest as biological change; shifts in the demersal fish communities are characterized by lags from the climate/ocean regime shift (Conners et al., 2002) and buildup of demersal fish stocks can exert top down control on populations of commercially important fish well after the climate shift (Bailey, 2000).
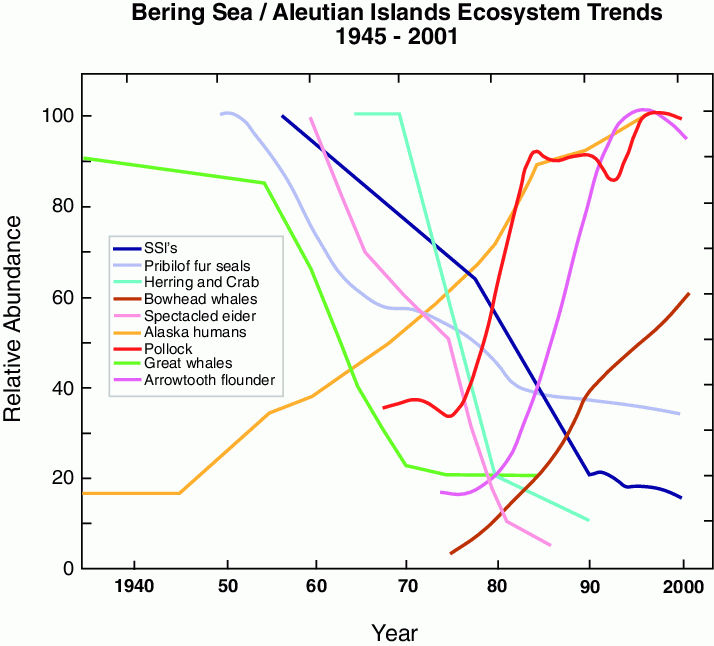
Figure 30.5: Schematic of ecosystem changes in the eastern Bering Sea showing trends of the relative abundance of select marine mammals, fish, and shellfish in the eastern Bering Sea (after NRC, 2003).
In the mid to late 1990s, the Eastern Bering Sea exhibited a host of noteworthy changes in physical and biological conditions (Kruse, 1998; Vance et al., 1998; Tynan, 1998; Baduini et al., 2001; Napp and Hunt, 2001; Hunt et al., 1999; Stabeno et al., 2001; Stabeno and Overland, 2001; Macklin et al., 2002; Napp et al., 2002; Olson and Strom, 2002): the first recorded major Bering Sea coccolithophorid blooms (1997, followed by blooms in 1998 through 2001), a large die-off of shearwaters (1997), salmon returns far below predictions (1997 and 1998), unusually warm summer sea surface temperatures (1997), unusually high ocean heat content (1998), and a decrease in the onshore transport of slope water (1997). While in the eastern Bering Sea, the abundance of pollock decreased (but continued to dominate biomass), a substantial increase occurred in gelatinous zooplankton (Brodeur et al., 1999a, 2002), arrowtooth flounder and other flatfish (Wilderbuer et al., 2002). Further, populations of Steller sea lions have continued their 30-year decline throughout the eastern Bering Sea. Below we address processes where the ocean temperature is directly related to sea ice processes and distribution, and to either wind or tidally generated turbulence and transport.
The pathways model (Fig. 30.2) considers sea ice as an important parameter, particularly fluctuations in extent, time of advance and subsequent melt-back, which are among the most striking physical phenomena (Schumacher and Stabeno, 1998) that also have profound impacts on biota (e.g., Hunt et al., 2002; Schumacher et al., 2003). Sea ice also provides habitat (e.g., haul outs) for some marine mammals, and once adequate solar radiation is available (early March), an under ice phytoplankton bloom occurs (Stabeno et al., 2001). The recent Oscillating Control Hypothesis (OCH; Hunt et al., 2002) relates climate change to energy flow and the timing of the plankton blooms, i.e., the timing of sea ice presence and melt-back is a critical aspect.
The interrelationships among the presence of sea ice, water column temperature, stability, and phytoplankton blooms (Fig. 30.6) have been observed from a long-term monitoring site on the middle shelf of the southeastern Bering Sea (Stabeno et al., 2001; Hunt et al., 2002). When ice is present in or after mid March, a strong peak occurs in chlorophyll fluorescence under the ice (e.g., 1995, 1997). The level of incoming short wave radiation prior to this time is likely insufficient to initiate a bloom of most phytoplankton species. The ice itself is relatively "thin" first year ice (<1 m) and has very little snow cover to attenuate or reflect incident radiation. When there is no ice present at this monitoring site (e.g., 2001), or the ice retreats before mid March (e.g., 1998, 2000), the bloom occurs in May or June. In 1999, sea ice was present at the site in late March, and returned in early May. As a result, there was an initial bloom in late March and another weaker and prolonged period of elevated fluorescence in late May and June.
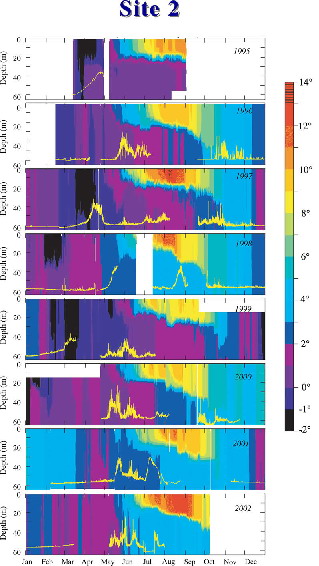
Figure 30.6: Time series of temperature contours at the long-term monitoring site in the middle domain (Site 2). Areas of black indicate cold water resulting from the presence of melting sea ice. The yellow line near the bottom of each panel indicates fluorescence at 11–13 m depth. For each year, the fluorometer tracts have been scaled to the highest value in that year. Gaps in this record occur due to fouling of the instrument or loss of mooring. (After Stabeno and Hunt, 2002.)
The spring bloom of phytoplankton associated with the sea ice accounts for 10–65% of the total annual primary production (Niebauer et al., 1995). A large proportion of the primary production from the sea-ice associated bloom eventually falls, unused, to the sea floor. This occurs because cold temperatures likely impact the ability of micro- and mesozooplankton grazers to effectively utilize the increase in production. Estimated total zooplankton production in 1999, a cold year, was 8–52% that of the two previous years when winter temperatures were average to above average (Coyle and Pinchuk, 2002a). While the total zooplankton production is dominated by small species that respond positively to temperature, at least one key species over the middle domain (Calayus marshallae) has higher standing stocks in colder years (Baier and Napp, 2003). In years when sea ice either does not exist over the southeastern shelf or retreats prior to the time when adequate light exists for a bloom (mid March), the spring bloom is delayed until later in the season when thermal stratification of the water column occurs (Stabeno et al., 2001). Under this scenario, phytoplankton biomass accumulates in the water column, but a much larger fraction of the primary production is utilized by zooplankton. The frequency of alternation between cold and warm regimes and the lengths of the individual stanzas may determine whether higher trophic levels are predominantly controlled by bottom-up or top-down mechanisms (Hunt et al., 2002; Hunt and Stabeno, 2002). In addition to how ice effects bottom up control of energy flow in the pelagic ecosystem, ice and its attendant cold pool directly influence the spatial distributions of higher trophic level biota (Ohtani and Azumaya, 1995; Wyllie-Echeverria and Ohtani, 1999; Brodeur et al., 1999b). As previously mentioned, avoidance of the cold pool often has the affect of increasing predation and cannibalism on larval and juvenile fishes by increasing the spatial overlap (or separation) of predator and prey.
In the Bering Sea, wind stress is another important physical force that strongly influences biological production as well as the flux of heat and nutrients (Fig. 30.2). Wind mixing largely determines the timing of the spring phytoplankton bloom when ice is not present, and helps to set annual new production through controlling the resupply of nitrate into the euphotic zone during summer (Sambrotto et al., 1986). Over the southeast shelf, the effect of wind mixing (through changes in water column stability) on nitrate uptake (primary production) can either decrease uptake during the respiration-limited phase or increase nitrate uptake once nutrients are limiting through resupply from deeper waters (Sambrotto et al., 1986). Not included in the pathway model are tides that strongly influence the southeastern Bering Sea. These two sources of turbulence are important to the region's biology and sediment processes.
The Inner Front Study, which began in 1997, focused on the importance of physical processes of the inner front to prolonged primary production. Results from Kachel et al. (2002) not only enhanced our knowledge of the inner front's physical characteristics, but they also elucidated some of its biological importance. During summer, nutrients could be pumped into the euphotic zone at the front, thereby stimulating production. The effectiveness of this process depends on two factors. First, a deep (sub-pycnocline) reservoir of nutrients must exist, and this is usually found within the middle shelf's bottom layer, the cold/cool pool. Typically, high concentrations of nutrients exist in the bottom layer over the middle shelf. An exception was 1997, when two factors (late spring storm and a shallow summer mixed layer) conspired to deplete nutrients from the cold pool. The strong, late spring storm vertically mixed the water column to a depth >50 m. Since nutrients in the euphotic zone had been depleted by an earlier, ice-associated, phytoplankton bloom, this caused the nutrient concentration in the cold pool to be reduced by almost half. Next, the very shallow mixed layer during the summer, allowed a phytoplankton bloom to occur beneath the surface mixed layer, which slowly consumed the nutrients in the cold pool (Stabeno et al., 2001; Stockwell et al., 2001). In addition to nutrients being available, sufficient mixing energy must occur to erode the extant vertical stratification. This can happen either by intensification of wind mixing and/or tidal mixing (throughout the fortnightly cycle); both of these processes move the inner front seaward. When this occurs, nutrients are made available to the frontal region, and are mixed upward where they can be utilized by phytoplankton. An example of this process can be seen in temperature and nitrate data collected across the inner front (Fig. 30.7). High concentrations occur within the cold/cool pool while those in the upper layer of the middle shelf and in the coastal domain are low. At the inner front, vertical finger-like structures with elevated concentrations of nitrate coincide with the 5.5°C isotherm. This structure was observed several days after an intense (wind speeds >14 m s![]() ) storm passed through the region and tidal currents were near their fortnightly maxima (Kachel et al., 2002).
) storm passed through the region and tidal currents were near their fortnightly maxima (Kachel et al., 2002).
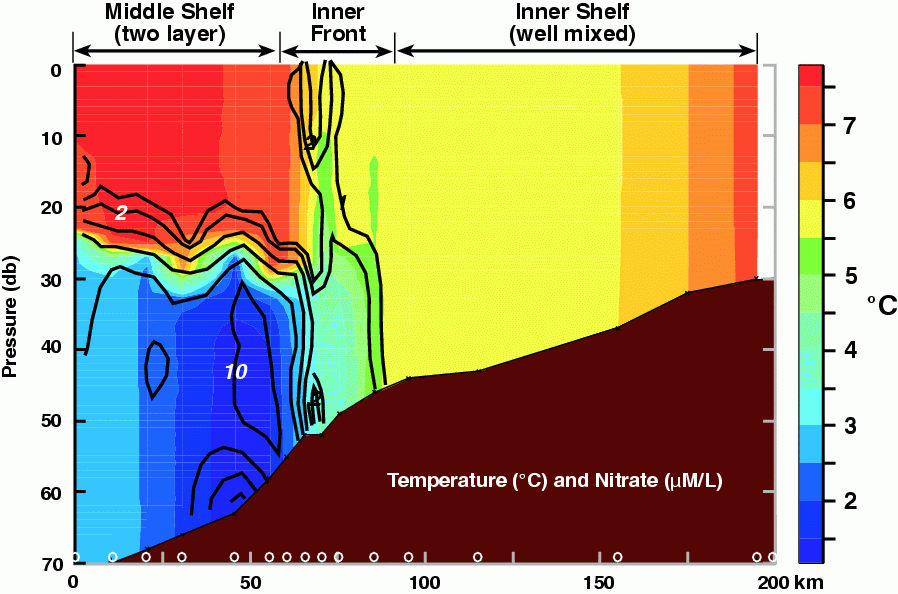
Figure 30.7: Contours of temperature (color; .C) and nitrate (black lines; mmoles l.1). The locations of the CTD stations are marked along the bottom axis. Nutrients were measured on each cast. (From Kachel et al., in 2002)
Turbulence also can directly affect the efficiency of larval feeding. Megrey and Hinckley (2001) used a process-oriented individual based model (IBM) of larval pollock that incorporated a turbulence-contact rate-feeding success mechanism, thus relating wind generated turbulence to feeding. Output from the model agreed with hydrodynamic theory, with a well-defined peak in consumption at intermediate wind speeds (MacKenzie et al., 1994). The functional form of wind speed versus maximum consumption was determined by a quadratic fit to model results. Optimum feeding (540 μg dry weight per day per individual) occurred at a wind speed of ~7 m s![]() . At wind speeds >9.5 m s
. At wind speeds >9.5 m s![]() increased turbulence negatively affected feeding, and at wind speeds <4.8 m s
increased turbulence negatively affected feeding, and at wind speeds <4.8 m s![]() feeding was less than optimal because turbulence was not sufficient to enhance contact rate. Fish and plankton can regulate their exposure to turbulence by adjusting their vertical position in the water column (Incze et al., 2001; Olla and Davis, 1990). In the Gulf of Alaska, survival of larval pollock cohorts has been tied to coincidence of the critical first feeding period and calm weather (Bailey and Macklin, 1994).
feeding was less than optimal because turbulence was not sufficient to enhance contact rate. Fish and plankton can regulate their exposure to turbulence by adjusting their vertical position in the water column (Incze et al., 2001; Olla and Davis, 1990). In the Gulf of Alaska, survival of larval pollock cohorts has been tied to coincidence of the critical first feeding period and calm weather (Bailey and Macklin, 1994).
Transport, which is indicated in the pathway model as changes in horizontal flow, is critical to many aspects of ecosystem dynamics, including advection of nutrients and plankton. Several studies suggest a connection between changes in climate and changes in recruitment of fish and shellfish species (e.g., Incze et al., 1987; Rosenkranz et al., 2001;Wilderbuer et al., 2002). One mechanism often cited is a change in the direction of transport of the planktonic stages (towards or away from favorable habitat or predators). Wespestad et al. (2000) related changes in climate and their regional impact on wind fields and transport of larval pollock in an attempt to identify sources of recruitment variability.
Recent studies have shown this shelf has considerable north-south variability. The southeastern shelf contains three distinct regions or latitudinal zones along the 70-m isobath: a strong two-layered system with cold/cool pool that lies south of about 57°N, an intermediate zone consisting of more well-mixed water; and a two-layered system with the northern cold pool that extends northward from about 58°N (Stabeno et al., 2002). In the bottom layer over the middle shelf, distributions of salinity and nutrients (nitrate) appear correlated. The greatest salinities and nitrate concentrations occurred in the northernmost zone and may represent the impact of across-shelf transport due to the mean flow in this region (Reed, 1998). Slope/outer shelf domain plankton taxa have been observed near the inner front as further evidence of this cross-shelf transport (Coyle and Pinchuk, 2002b).
Transport has important ramifications for dissolved and planktonic material, including larval fish (Wespestad et al., 2000; Wilderbuer et al., 2002) and crabs (Rosenkranz et al., 2001). Wespestad et al. (2000) used a simple wind drift model to generate trajectories of pollock eggs and larvae over the southeastern shelf. After a time period related to larval development, the young pollock were subjected to cannibalism by age-2 and older adults. While this approach was consistent with some year-classes of high recruitment, it did not fit the recruitment every year. One caveat is that the wind drift model only applies to the upper few meters of the water column, and both pollock eggs (Kendall, 2001) and larvae exist deeper in the water column (Napp et al., 2000). In addition, using a single initial point for the eggs/larvae is an oversimplification of the actual spawning regions for pollock (Hinckley, 1987).
To address these disparities, a different transport model was used to simulate transport. The North-eastern Pacific Regional Ocean Model System (NEPROMS) was selected to simulate drifter trajectories that more closely simulate pollock egg and larval transport (D. Ridgi, personal communications). The simulated pollock eggs were initialized in a grid, which contained the initial point used by Wespestad et al. (2000), but the drifter initial positions are denser near the surface, replicating egg distribution data collected in the Bering Sea (Kendall, 2001). A prominent feature in this region (north and east of Unimak Pass) is the confluence of flow of the Alaska Coastal Current into the Bering Sea through Unimak Pass and the upslope flow of the ANSC in Bering Canyon (Stabeno et al., 2002). The initial drifter positions were a seven by seven grid with horizontal separations of about 10 km at the confluence of two flows. Vertically, there were 15 drifters initialized at each grid point to a maximum depth just over 40 m. Drifters were released on April 1 of each year and tracked for 90 days.
Simulations were conducted for the period 1997–2001. In all years there was a tendency for drifters to move either toward the northeast along the Alaska Peninsula, or toward the northwest along the 100 m isobath. This result is in agreement with a schematic of mean circulation (Fig. 30.4) and satellite tracked drifter trajectories (Schumacher and Stabeno, 1998). Recently, abundances of age-0 pollock in and around the inner front at Cape Newenham were estimated to be the same order of magnitude as those around the Pribilof Islands (Coyle and Pinchuk, 2002b). Model trajectories of drifters from 1997 show that a subset of the drifters that began by following the 100 m isobath and then veered to the northeast. In 2000, trajectories revealed a strong turning to the northwest of trajectories that had been moving along the Alaska Peninsula. Fig. 30.8 shows trajectories for 2001. The simulations also suggest the importance of interannual variations in transport. They also suggest how dependent the end points of the "larval drift" are to small variations in both horizontal (order 10 km) and vertical (order 5–10 m) initial positions. Numerous trajectories generated by a model, which includes only wind drift (OSCURS: Wespestad et al., 2000) were toward the northeast along the Alaskan Peninsula, as were the majority of the NEPROMS simulations in the upper five divergence of the trajectories. In the 5–20 m and 20–40 m release bins there were many drifters that followed the 100 m isobath to the northwest, with some even moving through Unimak Pass into the North Pacific Ocean before turning back. Further examination is required to determine the environmental parameters that resulted in the interannual differences in trajectories, and therefore to help understand how transport affects year class strength of pollock and other plankton.
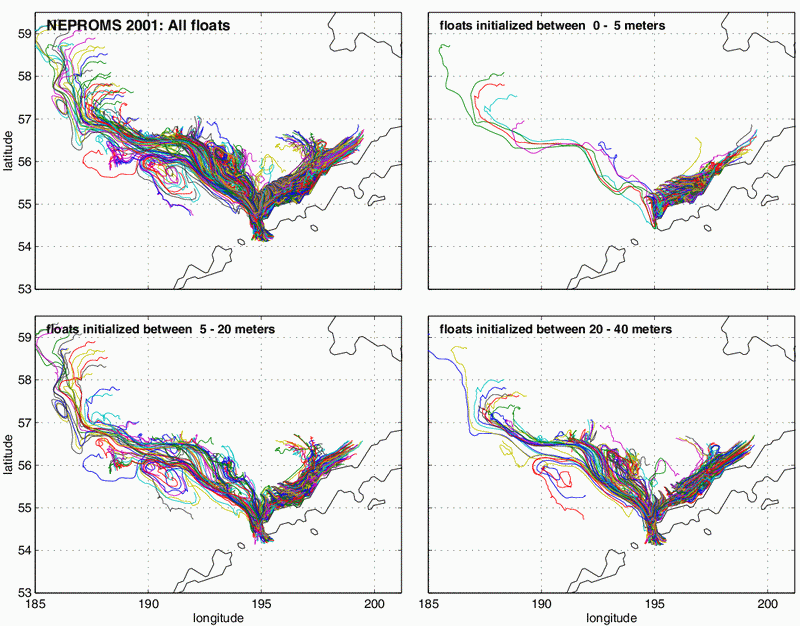
Figure 30.8: Trajectories simulated for a 90-day period starting on 1 April 2001. The upper left panel shows all drifters, while the other panels show drifters divided as a function of initial release depth. Note that with increasing depth there is a greater tendency for drifters to flow northwestward and even become involved in the eddy-like circulation associated with the Bering Slope Current. This suggests a mechanism for planktonic transport between oceanic and shelf waters.
While many of the observations of recent changes come from the southeastern shelf, similar ecosystem-level change has been observed in the northern Bering Sea. There are indications of changes in benthic biomass south of St. Lawrence Island, and in the Chirikov Basin between Lawrence Island and Bering Strait (Grebmeier and Cooper, 2002). Studies begun in the mid-1980s have shown declines in the biomass (Grebmeier, 1993; Sirenko and Koltun, 1992; Grebmeier and Dunton, 2000) and mean sizes of the dominant bivalves in the area (Grebmeier and Cooper, 2002). Sediment respiration rates, which indicate carbon loading to the sea floor, have also declined since the late 1980s. Seasonal patterns of sediment chlorophyll concentrations show that deposition of carbon in this area is related to the ice-edge spring bloom (Cooper et al., 2002), thus any change in the timing of ice retreat during the late winter/early spring will likely have a major impact on this ecosystem. Although commercial fishing may have played a role in changes to the southern Bering Sea ecosystem, there is little commercial fishing on the northern shelf where changes in benthic faunal populations and declines in dominant fauna have occurred, resulting in a cascade effect on higher trophic levels (Grebmeier and Dunton, 2000; Grebmeier and Cooper, 2002).
Ecosystem processes related to sea ice are dominant features of the northern shelf. In years with extreme occurrences of sea ice (e.g., extremely high in 1975/76 and extremely light in 1978/79), sea ice can be present at 62°N over the northern shelf from mid-November until mid-June, or only from late January to late April. While the presence of sea ice provides a substrate for marine mammals, it also limits at-sea observations. In other regions, under-ice algae provide a wintertime food source for zooplankton during periods of reduced water column production (Tourangeau and Runge, 1991). Most of our knowledge of this high latitude ecosystem comes from summertime observations. Primary production here is not consumed by pelagic secondary consumers (Coyle and Cooney, 1988; Springer and McRoy, 1993), but rather by a rich macro-benthic community. Large populations of benthic, rather than pelagic-feeding marine mammals and birds serve as apex predators in this food chain (Grebmeier and Cooper, 1995).
The primary physical process that results in high primary production over the northern shelf is the onshelf transport of nutrient-rich waters, which are then advected across the shelf. This current, known as the Anadyr Current (Shuert and Walsh, 1993), flows through Anadyr Strait and then northward through Bering Strait (Schumacher and Stabeno, 1998). As the bathymetry shoals (<40 m) in the northern Gulf of Anadyr, nutrients enter the euphotic zone and a production-deposition center is formed (Coachman, 1993). Another center is located over the Chirikov basin north of St. Lawrence Island. As the Anadyr Current flows between St. Lawrence Island and Siberia, the bathymetry deepens (50–60 m) and bottom-generated turbulence results in strong vertical mixing. This tends to mix phytoplankton below the critical depth and interrupts the bloom in the strait (Springer and McRoy, 1993). As the current speed is reduced over Chirikov Basin, a region of extremely high primary production occurs (12–16 gCm![]() day
day![]() ; Springer and McRoy, 1993). The strength of the Anadyr Current is related to the magnitude of flow through Bering Strait. This net northward flow is driven by the difference between sea level in the North Pacific and Arctic Oceans (0.4–0.5 m). On short time scales, this northward flow is modified by wind-generated coastal changes in sea level (Aagaard et al., 1985; Overland and Roach, 1987). During weak winds of summer, northward transport is greatest (Coachman, 1993) and provides the strong flux of nutrients for primary production. The Anadyr Current also transports a high biomass of large oceanic copepods from the slope regions of the basin onto the northern shelf (Springer et al., 1989). In the Chirikov Basin, these support high numbers of planktivorous auklets (Springer and Roseneau, 1985; Springer et al., 1987) that forage in stratified Bering shelf water (Hunt et al., 1990) or in frontal regions on aggregations of these copepods (Hunt and Harrison, 1990). This combination of biophysical coupling supports a pelagic seabird community far from the origin of the zooplankton on which they are dependent.
; Springer and McRoy, 1993). The strength of the Anadyr Current is related to the magnitude of flow through Bering Strait. This net northward flow is driven by the difference between sea level in the North Pacific and Arctic Oceans (0.4–0.5 m). On short time scales, this northward flow is modified by wind-generated coastal changes in sea level (Aagaard et al., 1985; Overland and Roach, 1987). During weak winds of summer, northward transport is greatest (Coachman, 1993) and provides the strong flux of nutrients for primary production. The Anadyr Current also transports a high biomass of large oceanic copepods from the slope regions of the basin onto the northern shelf (Springer et al., 1989). In the Chirikov Basin, these support high numbers of planktivorous auklets (Springer and Roseneau, 1985; Springer et al., 1987) that forage in stratified Bering shelf water (Hunt et al., 1990) or in frontal regions on aggregations of these copepods (Hunt and Harrison, 1990). This combination of biophysical coupling supports a pelagic seabird community far from the origin of the zooplankton on which they are dependent.
Flow along the Alaskan coast of the northern Bering Sea is a combination of river runoff (mainly the Yukon River) and a continuation of the BCC of the southeastern Bering Sea (Schumacher and Stabeno, 1998). Production here has been identified as typical of shallow shelves elsewhere: once nutrients are exhausted during an initial bloom, production is low and only ~10% of that for Chirikov Basin and the Gulf of Anadyr (Springer and McRoy, 1993). The east-west gradient in water properties, nutrients, primary production and fauna (transported in the Anadyr Current) result in two different community structures, a rich benthic system to the west and a less productive eastern system (McRoy, 1993). Benthic macro faunal biomass ranges from ~30–60 gCm![]() for the western portion and <10 gCm
for the western portion and <10 gCm![]() in the Alaskan coastal waters.
in the Alaskan coastal waters.
Over the northern shelf, regions exist that are usually ice-free throughout wintertime, and these regions are known as polynyas. These features often are found on down-wind facing coasts (e.g., the south coast of St. Lawrence Island). In the polynyas ice formation is caused by frigid winds blowing from the northeast. This and the resulting brine rejection are important features of the physical environment. As ice is formed, brine is released and sinks to the bottom. The current field generated by this process results in both offshore flow and sometimes a reversal of the mean eastward flow of the branch of the Anadyr Current that flows south of St. Lawrence Island (Schumacher et al., 1983). This wintertime set of physical processes apparently affects sediment patterns, e.g., surface C/N ratios, total organic carbon, sediment oxygen uptake, and benthic biomass (Grebmeier and Cooper, 1995). As a result of the enhanced production, the region southwest of St. Lawrence Island has long been a favorite feeding ground for gray whales.
Recent changes have been observed in the ecosystem of the western Bering Sea (Radchenko et al., 2001). Although the Bering Sea responds to climate shifts, a significant portion of the variability occurs on year-to-year time scales, which can make the identification of regime shifts difficult. This year-to-year variability is clearly evident in recent years. For instance, 1999 was an unusually cold year, characterized by negative anomalies in atmosphere and sea surface temperatures, and extensive and prolonged sea ice cover. In contrast, positive sea surface temperature anomalies were predominant throughout the northwestern shelf of the Bering Sea in 1997 (spring and summer) and 1998 (summer). In response to the physical conditions in 1999, some biological processes were less active and/or delayed. During 1999, the return of Pacific salmon to streams in northeastern Kamchatka was approximately two weeks later than typical.
Over the continental shelf of the western Bering Sea, the importance of transport to distribution of gonatid squid (Berryteuthis magister) has recently been investigated (Arkhipkin et al., 1998). This squid is one of the most abundant in the North Pacific Ocean/Bering Sea and is exploited commercially by both Russian and Japanese bottom trawl fisheries. Distinct patterns of size composition, age structure and growth occur over the western shelf, and given the life history of this squid, time-dependent transport of planktonic stages provides the most likely explanation (Arkhipkin et al., 1998). While much of the counter-clockwise current over the Bering Sea basin is related to inflow of the Alaskan Stream through passes in the Aleutian Island Chain (Overland et al., 1994; Stabeno et al., 1999; Stabeno et al., 2004), seasonal changes in wind stress can alter transport in the gyre by ~50% (Bond et al., 1994). As a result of this strong seasonal signal, regional shelf edge current patterns in the western Bering Sea apparently have two modes: a strong along slope mode and an eddy mode that exists as the current relaxes (Verkhunov and Tkachenko, 1992). As noted in Section 3.4, this is also the case for the eastern BSC. Associated with these modes are two primary transport routes for the juvenile squid: winter and spring hatched juveniles are first transported into the eastern region of the shelf and then westward along the shelf; spring/summer hatched juveniles are transported directly across the basin into the western region of the shelf (Verkhunov and Tkachenko, 1992).
In an analysis of long-term fluctuations in several species of pelagic and benthic fishes of the western Bering Sea, Naumenko (1996) notes the correspondence between changes in fish biomass and conditions in the ecosystem. Using catch data, he discerned that the fish community could be divided into four periods: 1958–1964, when herring dominated, 1965–1974, a transitional period when no single species dominated and stocks were highly variable, 1975–1987, when pollock dominated in both biomass and abundance, and 1987–1993 (last data set presented), when pollock declined and groundfish populations increased. While some of the fluctuations could be accounted for by fishing mortality, it appeared that environmental conditions have played an important role in the fluctuations. The first connection Naumenko (1996) made was between bottom water temperatures in fall and surface temperatures in spring and summer, with the four fish community periods. The first period was moderately warm, the second abnormally cold, the third abnormally warm. The fourth period appeared to be one of transition. He notes that changes in the biomass of zooplankton closely correlated with the temperature changes, with zooplankton biomass being greatest during the warm period and lowest in the cold period. Note the correspondence of these observations to the OCH (discussed in section 7): cold regime implies low zooplankton biomass and warm regime high biomass. Changes in recruitment of pollock in the western Bering Sea have also been attributed to changes in the climate/oceanic regime (Balykin, 1996), although the processes that might link the climate/oceanic system to fish survival were not discussed.
Efforts to model the eastern Bering Sea ecosystem were reviewed by Francis et al. (1999). Models of ecosystem dynamics of the southeastern Bering Sea shelf span the full spectrum from conceptual models (e.g., the OCH, Hunt et al., 2002; the Trophic Cascade Hypothesis, Merrick, 1995; NRC 1996), to trophic or food web models (e.g., Laevastu and Larkins, 1981; Trites et al., 1999), to models that explicitly include links between specific ecological processes, such as physical processes and primary/secondary production (Hood, 1999; Walsh and McRoy, 1986). After providing some essential background information on the ecosystem, we focus on the OCH conceptual model. This model both encompasses most of the previous results and ideas regarding how the ecosystem of the southeastern Bering Sea functions, and it extends the conceptual linkages of processes, which were initially developed in the PROBES study (Walsh and McRoy, 1986).
Currently, pollock is the most abundant species harvested in the Bering Sea, accounting for >65% of the total groundfish biomass; during the 1980s their total biomass exceeded 20 million metric tons (Napp et al., 2000). The biomass trends in the eastern Bering Sea (1979 to 1998: Schumacher et al., 2003) show that while the total biomass of pollock in the 1990s was less than in the 1980s, they still dominate biomass in any of the trophic guilds which include marine birds, mammals, other fishes and crabs. Walleye pollock is a nodal species in the food web (NRC Report, 1996) with juveniles being the dominant prey of fishes (including adult pollock), seabirds, and marine mammals (Springer and Byrd, 1989; Livingston, 1993). The abundance of pollock is determined by episodic occurrence of strong year-classes. Such a strong year-class occurred in 1978, the remaining year-classes fluctuating about the long-term mean abundance (Fig. 30.9).
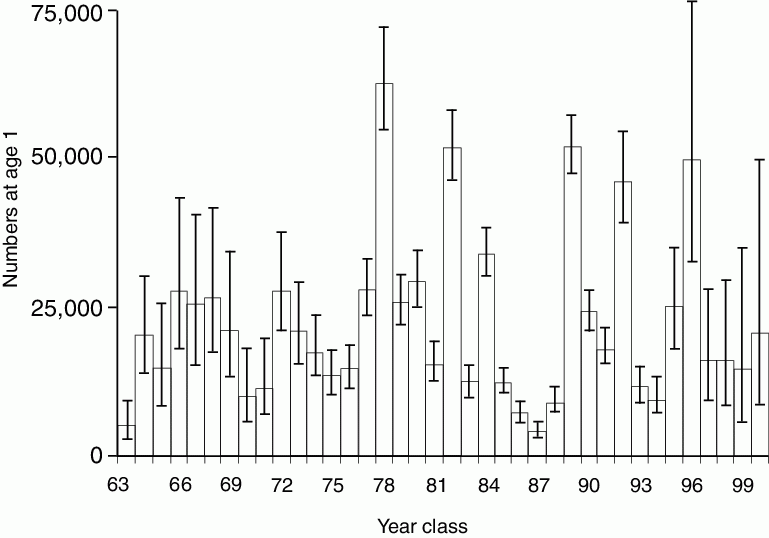
Figure 30.9: Time series of year class abundance of age-1 pollock. Error bars represent ~95% confidence interval. (From Ianelli et al., 2001.)
The observational basis for the OCH includes many of the elements of the pathway model, including sea ice, wind generated turbulence, water column temperature and chlorophyll fluorescence (Fig. 30.10). The observations of biota included zooplankton, jellyfish, and four species of fish, marine mammals and sea birds. After an analysis of these data, Hunt et al. (2002) formulated the OCH. The OCH recognizes that late retreat of sea ice with the attendant cold temperatures in the water column and early, short phytoplankton blooms are hallmarks of cold regimes. The associated impacts on biota include reduced survival of fish eggs (Blood, 2002) and diminished production of zooplankton prey for larval fish. Under this set of conditions, the hypothesis predicts that recruitment of pollock will be nominal or weak, and strong year-classes are not expected. Bottom-up processes dictate the flow of energy through the ecosystem during a cold regime (Fig. 30.11). Cold water-column temperatures can directly impact distributions of some forage fish species. The OCH allows that pinnipeds and piscivorous seabirds may thrive, even under cold conditions, if the population centers of forage fish change and become more available as prey. During years when sea ice is either not present or retreats before there is sufficient net short wave radiation to initiate a bloom, the spring bloom occurs later than during the cold regime, and water column temperatures are warmer. Under this set of conditions, the spring bloom is expected to be prolonged and zooplankton production is expected to be strong, resulting in readily available prey for larval and juvenile fish. The potential then would be high for strong year-classes of pollock and other piscivorous fish (Fig. 30.11).
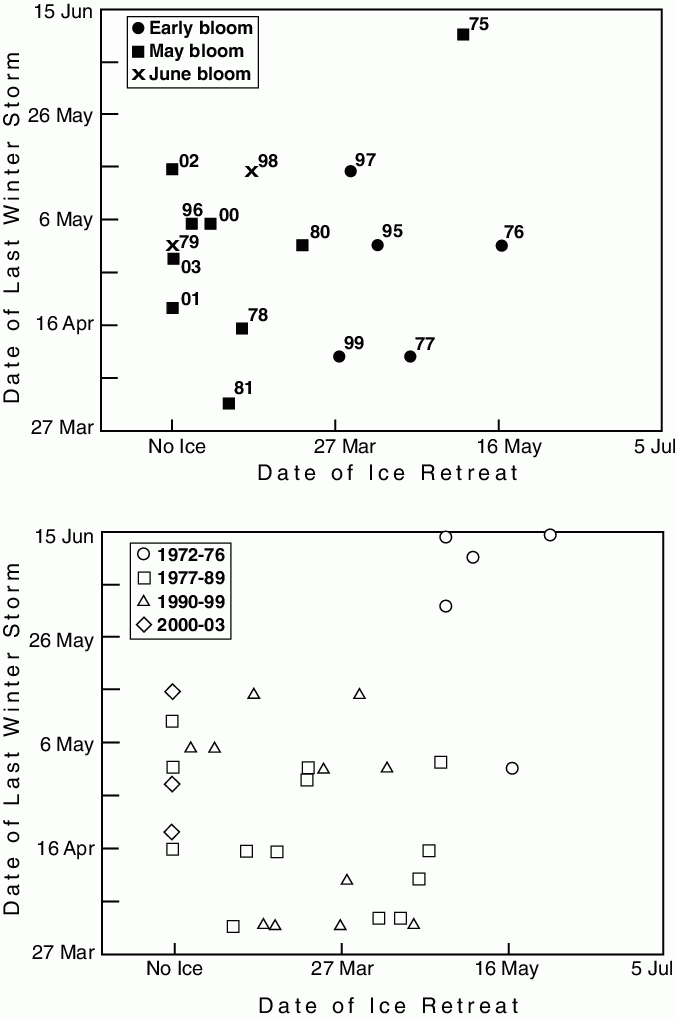
Figure 30.10: A synthesis of physical phenomena that help dictate the timing of the spring phytoplankton bloom of the southeastern Bering Sea shelf. The top panel includes the timing of the last retreat of sea ice, the timing of the shift between winter and summers wind conditions, and their effect on the timing of the spring bloom over the southern portion of the middle domain. Note that ice-related or early blooms occur in years when the ice retreat comes in mid March or later. In 1975 there was an ice-related bloom in May. The date of the last winter storm was defined as when the wind speed cubed fell below 2500 m![]() s
s![]() for the summer. The winds were measured at St. Paul Island. (From Hunt et al., 2002.) The lower panel shows the relationship between ice retreat and date of last winter storm with the years divided into regimes. If the relationship between timing of ice retreat and spring phytoplankton bloom hold, then the timing of the spring bloom can be inferred for each regime.
for the summer. The winds were measured at St. Paul Island. (From Hunt et al., 2002.) The lower panel shows the relationship between ice retreat and date of last winter storm with the years divided into regimes. If the relationship between timing of ice retreat and spring phytoplankton bloom hold, then the timing of the spring bloom can be inferred for each regime.
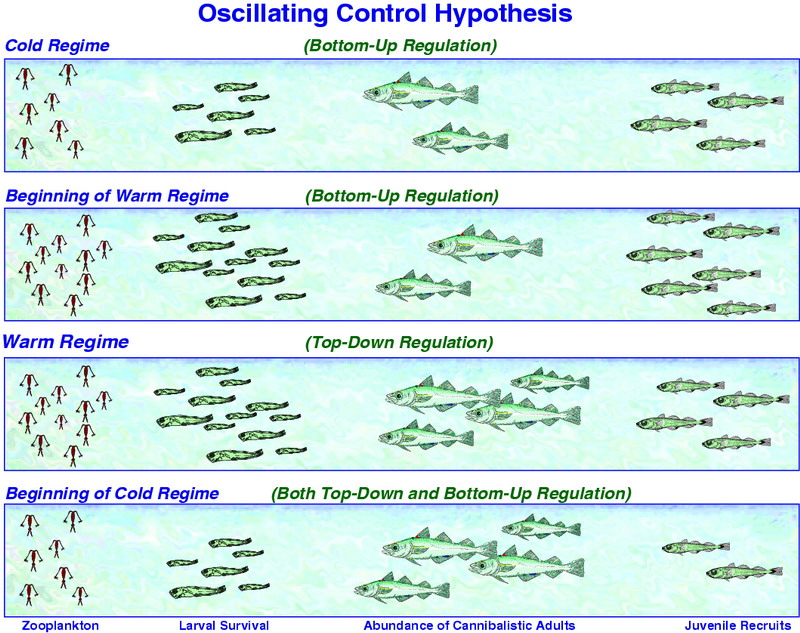
Figure 30.11: Schematic of the OCH. The number of pollock recruits affects not only pollock population dynamics, but also the availability of age-1 pollock to predators such as marine birds and mammals. Top Panel: In a cold regime, copepod production is limited by water temperature and larval/juvenile pollock survival is prey limited. Upper Middle Panel: At the beginning of a warm regime, copepods flourish and provide ample food resources to support strong survival of larval and juvenile pollock; juvenile pollock survival is high because there are few large piscivorous fish to consume them. Lower Middle Panel: In a warm regime, the production of copepod food for larval and juvenile pollock remains strong, but survival of juveniles and recruitment is limited to cannibalism from the growing biomass of adult pollock and other predators such as Pacific cod and arrowtooth flounder. Bottom Panel: At the beginning of a cold regime, copepod production is limited and larval and juvenile pollock are food limited. In addition, cannibalism by adult pollock and other predators further limits recruitment. Thus, there are asymmetries in how pollock recruitment will respond to changes from a cold to a warm regime versus from a warm to a cold regime.
There are years when the observed regime conditions (e.g., 1976 – cold with low recruitment, 1996 – warm with high recruitment) and estimated pollock production fit the outcomes predicted by the OCH; however, there are also years when the fit is poor (e.g., 1987 – warm with low recruitment, 1992 – cold with high recruitment). The history of the predator field (i.e., top-down process) on young pollock, therefore, must also be taken into account. The marked impact of adult pollock on age-1 and younger fish, together with other fish (e.g., arrowtooth flounder and Pacific cod) is well documented (Livingston et al., 1999; Livingston and Methot, 1998). The OCH weaves in such biological mechanisms as follows. When there is a sequence of warm regime years, recruitment is above average and the populations of adult predatory fish will eventually increase to a point where the control of future year-class strength is mainly a top-down process. This switch to top-down dominance may have occurred for the pollock population in the Gulf of Alaska (Bailey, 2000). There, as the arrowtooth flounder population markedly increased, the pollock population underwent a severe decline and the "critical period" for determining year-class strength switched from the egg and larval stages to the juvenile period. The hypothesis predicts that as predation becomes greater, the abundance of young pollock and forage fish decline and zooplankton become available for other populations (e.g., jellyfish, salmon, baleen whales). In addition, the reduction in abundance of forage fish could cause declines in populations and/or productivity of pinnipeds and piscivorous seabirds (Hunt and Stabeno, 2002).
While decadal time scale or regime shifts have been identified for the Bering Sea (e.g., Stabeno et al., 2001; Minobe, 2002; Hare and Mantua, 2000), much of the variance of physical variables lies in the interannual frequency band. The OCH addresses the impact of annual changes in the following manner. When a cold year occurs in a warm regime, the inertia of the large biomass of predatory adult fish maintains a top-down energy flow. If sufficiently large year-classes of piscivorous predators (e.g., pollock) result from one or two warm years in a cold regime, the control could switch from bottom-up to top-down. Variations in forage fish and pollock year-class strength can also occur within a regime as a result of the redistribution of their predators.
A substantial amount of information has been compiled which identifies detailed needs for future interdisciplinary research in coastal waters of the eastern Bering Sea (e.g., NRC, 1996; Draft Bering Sea Ecosystem Research Plan, 1998; Springer, 1999). Further, Hunt et al. (2002) present seven testable relationships among physical and biological processes that would lead toward substantiation or refutation of the OCH. Of these, five require comprehensive monitoring of biophysical features of the shelf.
At present, there are several monitoring efforts underway in the Bering Sea. Among the most notable are the annual trawl and acoustic surveys conducted by the National Marine Fisheries Service. The ensuing time series of water temperature, and biomass/abundance of many fish and other marine biota form the basis of numerous publications regarding the Bering Sea ecosystem (Ianelli et al., 2001). Bird (Dragoo et al., 2000) and marine mammal counts (National Marine Mammal Laboratory, Seattle, WA) are being conducted at many sites in the region and supplement the survey observations. Sea ice is well monitored by satellite observations; water column currents and properties, chemistry, and ichthyo-zooplankton populations, however, are greatly under-sampled. When we consider that the eastern shelf is approximately the same size as the states of California, Oregon, and Washington combined and that at present there is only one continuously occupied site (1995–present, Site 2 in the middle shelf of the southeastern Bering Sea) where biophysical moored instruments are maintained, it is amazing we have learned as much as we have. Recent re-analyses of long biological time series exemplify the great degree of inherent variability on the Bering Sea shelf that must be taken into account when attempting to design new monitoring studies (Napp et al., 2002). We note that the time series from Site 2 (Fig. 30.6) were a keystone in formulating the OCH and have formed the basis for more than a dozen publications and supportive data for many others. A more comprehensive, permanent monitoring program is needed for Alaskan coastal waters.
The realization that there is value to the traditional knowledge and wisdom (TKW) of the local Native Peoples has come slowly, but is now gaining recognition (NRC, 1996). Native People rely for their existence on their ability to see the patterns in all elements of their environment: the weather, sea ice, fish, marine mammals and birds. This knowledge is now finding its way into scientific literature (e.g., Pungowiyi, 2000; Noongwook, 2000; Huntington, 2000). While TKW is not quantitative, it has the potential to make contributions to both retrospective and current studies. Its use as an integrative tool will help us to better understand the ecosystem of the Bering Sea. Further, it is obvious that the Native Peoples could provide a valuable component for monitoring the vast Bering Sea (e.g., Harwood et al., 2002).
On the national level, the National Ocean Research Leadership Council (NORLC), a Cabinet-level group of 14 Federal agencies, has designated the establishment of the U.S. Integrated Ocean Observing System (IOOS) as its highest priority. The process was accelerated in 2000 with the creation of Ocean.US, a federal interagency office established by the National Oceanographic Partnership Program (NOPP). To date, nine agencies have agreed to participate in the Ocean.US endeavor. Establishment of an IOOS is driven by both national and regional (multi-state) priorities for data and informational products. Among the objectives of this proposed program are: to improve predictions of climate change and its effects on coastal communities, to protect more effectively and restore healthy coastal marine ecosystems, and to enable the sustained use of marine resources.
In addition to observations, expansion of the modeling effort is necessary to better understand dynamics of the Bering Sea ecosystem. Modeling can be portioned into two aspects. Hydrodynamic models can treat biological components (e.g., pollock eggs and larvae) as "floats" so that the model becomes an individual based model of ecosystem dynamics containing many of the elements of the conceptual pathway model (e.g.,Walsh and McRoy, 1986). Another approach is the application of the ECOSIM model (e.g., Walters et al., 1999; Aydin, 2002), which is a population-scale quantitative food web model that adheres to a mass-balance (not necessarily equilibrium), where the linkage to physics can enter though bottom-up forcing on primary production. Both approaches have value and both suffer because of the complex nature of using them as simple tools to examine biophysical processes and mechanisms. We believe that models and/or their informational products need to become more "user friendly" and generally available for all investigators.
How can the NEPROMS become a user-friendlier tool? The answer to this question could take the form of developing informational products from model simulations. Computed and averaged (perhaps on a daily time step) water property and velocity field distributions could be stored. Coupling these averaged fields with an interactive interface, which would allow a naive user to choose particular simulations and then access them in selected time/space domains, would permit far more research into questions of how the ecosystem functions than is possible today.
The Bering Sea supports a rich and productive ecosystem, with strong physical forcing and a well-defined climate signal. Economically, it is extremely valuable, providing nearly half of the US catch of fish and shellfish. It provides sustenance and the basis for important cultural traditions of native communities throughout the region. Although its economic importance has been acknowledged for some time, long-term biological and physical observations do not adequately cover the vast area of the Bering Sea. New technologies of satellite observation, moored autonomous samplers, and autonomous vehicles may help to solve the problems of under sampling. Multi-national research endeavors in this region would also help us to obtain sufficient sample densities to describe the mechanisms that structure this ecosystem. The Bering Sea ecosystem is ripe for description and improving our understanding of the biophysical mechanisms that lead to its high productivity.
Much of the material presented in the Chapter is taken from three National Science Foundation research programs (PROBES, ISHTAR and INNER FRONT), two National Oceanic and Atmospheric Administration (NOAA) Coastal Ocean Programs studies (Bering Sea Fisheries Oceanography Coordinated Investigations and Southeast Bering Sea Carrying Capacity), and from NOAA's ongoing Fisheries Oceanography Coordinated Investigations program conducted jointly by the Pacific Marine Environmental Laboratory and the Alaska Fisheries Science Center, both in Seattle, WA. J. Schumacher acknowledges funding from the PMEL/FOCI program for his time in writing this Chapter, and G. Hunt received support from NSF Grant OPP-9617287. NOAA's Coastal Ocean Program sponsored some of this research and this chapter is contribution FOCI-B451 to Fisheries-Oceanography Coordinated Investigations, and is contribution #2529 from PMEL.
Aagaard, K., L.K. Coachman, and E. Carmack, 1985. On the halocline of the Arctic Ocean. J. Geophys Res., 90, 4833–4846.
Alexander, V., 1999. Interdisciplinary studies of the Bering Sea. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), University of Alaska Sea Grant, AK-SG-99-03, 683–686.
Arkhipkin, A.I., V.A. Bizikov, and A.V. Verkhunov, 1998. Distribution and growth in juveniles of the squid Berryteuthis magister (Cephalopoda, Gonatidae) in the western Bering Sea. Sarsia, 83, 45–54.
Arsenev, V.S., 1967. Currents and water masses of the Bering Sea. Nuka, Moscow, 135 pp.
Aydin, K., 2002. The use of ecosystem models to investigate multispecies management strategies for capture fisheries, In: Fish. Cent. Res. Rep., T. Pitcher and K. Cochrane (eds.), 33–38.
Baduini, C.L., K.D. Hyrenbach, K.O. Coyle, A. Pinchuk, V. Mendenhall, and G.L. Hunt, Jr., 2001. Mass mortality of short-tailed shearwaters in the southeastern Bering Sea during summer 1997. Fish. Oceanogr., 10(1), 117–130.
Baier, C.T., and J.M. Napp, 2003. Climate-induced variability in Calanus marshallae populations. J. Plankton Res., 25, 771–782.
Bailey, K.M., 2000. Shifting control of recruitment of walleye pollock Theragra chalcogramma after a major climate and ecosystem shift. Mar. Ecol. Prog. Ser., 198, 215–224.
Bailey, K.M., and S.A. Macklin, 1994. Analysis of patterns in larval walleye pollock Theragra chalcogramma survival and wind mixing events in Shelikof Strait, Gulf of Alaska. Mar. Ecol. Prog. Ser., 113, 1–12.
Bailey, K.M., T.J. Quinn, W.S. Grant, and P. Bentzen, 1999. Population structure and dynamics of walleye pollock, Theragra chalcogramma. Adv. Mar. Biol., 37, 179–255.
Bailey, K.M., L. Ciannelli, N.A. Bond, A. Belgrano, and N.C. Stenseth, 2005. Recruitment of walleye pollock in a physically and biologically complex ecosystem: A new perspective. Prog. Oceanogr., 67, 24–42.
Balykin, 1996. Dynamics and abundance of Western Bering Sea Walleye pollock. In: Ecology of the Bering Sea: A review of Russian literature, Mathiesen, O.A. and K.O. Coyle (eds.), University of Alaska Sea Grant Program Report No. 96-01, Fairbanks, AK, 177–182.
Blood, D.M., 2002. Low-temperature incubation of walleye pollock (Theragra chalcogramma) eggs from the southeast Bering Sea shelf and Shelikof Strait, Gulf of Alaska. Deep-Sea Res. II, 49, 6095–6108.
Bond, N.A, J.E. Overland, and P. Turet, 1994. Spatial and temporal characteristics of the wind forcing of the Bering Sea. J. Climate, 7, 1119–1130.
Bond, N.A., J.E. Overland, M. Spillane, and P.J. Stabeno, 2003. Recent shifts in the state of the North Pacific. Geophys. Res. Lett., 30(23), 2183, doi: 10.1029/2003GL018597.
Brodeur, R.D., C.E. Mills, J.E. Overland, G.E. Walters, and J.D. Schumacher, 1999a. Evidence for a substantial increase in gelatinous zooplankton in the Bering Sea, with possible links to climate change. Fish. Oceanogr., 8, 296–306.
Brodeur, R.D , M.T. Wilson, G.E. Walters, and I.V. Melnikov, 1999b. Forage fishes in the Bering Sea: distribution, species associations and biomass trends. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 509–536.
Brodeur, R.D., H. Sugisaki, and G.L. Hunt, Jr., 2002. Increases in jellyfish biomass in the Bering Sea: Implications for the ecosystem. Mar. Ecol. Prog. Ser., 233, 89–103.
Coachman, L.K., 1986. Circulation, water masses, and fluxes on the southeastern Bering Sea shelf. Cont. Shelf Res., 5, 23–108.
Coachman, L.K., 1993. On the flow field in the Chirikov basin, Cont. Shelf Res., 13, 481–508.
Coachman L.K., K. Aagaard, and R.B. Tripp, 1975. Bering Strait: The regional physical oceanography. Univ. Wash. Press, Seattle, 172 pp.
Conners, M.E., A.B. Hollowed, and E. Brown, 2002. Retrospective analysis of Bering Sea bottom trawl surveys: regime shift and ecosystem reorganization. Prog. Oceanogr., 55, 209–222.
Cooney, R.T., and K.O. Coyle, 1982. Trophic implications of cross-shelf copepod distributions in the southeastern Bering Sea. Mar. Biol., 70, 187–196.
Cooper, L.W., J.M. Grebmeier, I.L. Larsen, V.G. Egorov, C. Theodorakis, H.P. Kelly, and J.R. Lovvorn, 2002. Seasonal variation in sedimentation of organic materials in the St. Lawrence Island polynya region, Bering Sea. Mar. Ecol. Prog. Ser., 226, 13–26.
Coyle, K.O., and R.T. Cooney, 1988. Estimating carbon fluxes to pelagic grazers in the ice-edge zone of the eastern Bering Sea. Mar. Biol., 98, 299–306.
Coyle, K.O., and A.I. Pinchuk, 2002a. Climate-related differences in zooplankton density and growth on the inner shelf of the southeastern Bering Sea. Prog. Oceanogr., 55, 177–194.
Coyle, K.O., and A.I. Pinchuk, 2002b. The abundance and distribution of euphausiids and zero-age pollock on the inner shelf of the southeast Bering Sea near the Inner Front in 1997–1999. Deep Sea Res. II, 49, 6009–6030.
Cury, P., C. Roy, R. Mendelssohn, A. Bakun, D.M. Husby, and R.H. Parrish, 1995. Moderate is better: exploring nonlinear climate effects on the Californian northern anchovy (Engraulis modax). Canadian Special Publication of Fisheries and Aquatic Sciences, 108, 417–424.
Draft Bering Sea Ecosystem Research Plan, 1998. Copies available from Pat Livingston, AFSC/ NMFS, Bldg. #4, 7600 Sand Point Way NE, Seattle WA, 98115-0070, 58 pp.
Dragoo, D.E., G.V. Byrd, and D.B. Irons, 2000. Breeding status and population trends of seabirds in Alaska in 1999. U.S. Fish and Wildl. Serv. Report AMNWR 2000/02, 61 pp.
Favorite, F., A.J. Dodimead, and K. Nasu, 1976. Oceanography of the subarctic Pacific region, 1960– 71. Int. N. Pac. Fish. Comm. Bull., 33, 187 pp.
Fay, F.H., 1981. Marine mammals of the Eastern Bering sea shelf: An overview. In: The Eastern Bering Sea Shelf: Oceanography and Resources, Vol. II, D.W. Hood and J.A. Calder (eds.), 807– 811.
Ford, C., 1966. Where the sea breaks its back: The epic story of early naturalist George Steller and the Russian exploration of Alaska. Alaska Northwest Books, Portland, OR, 206 pp.
Francis, R.C., S.R. Hare, A.B. Hollowed, and W.S. Wooster, 1998. Effects of interdecadal climate variability on the oceanic ecosystems of the NE Pacific. Fish. Oceanogr., 7, 1–21.
Francis, R.C., K. Aydin, R.L. Merrick, and S. Bollens, 1999. Modeling and management of the Bering Sea Ecosystem. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), University of Alaska Sea Grant, AK-SG-99-03, 409–433.
Giese, B.S., S.C. Urizar, and N. Fučkar, 2002. Southern Hemisphere origins of the 1976 regime shift. Geophys. Res. Lett., 29(2), 10.1029/2001GL013268.
Grebmeier, J.M., 1993. Studies of pelagic-benthic coupling extended into the Soviet continental shelf in the northern Bering and Chukchi Seas. Cont. Shelf Res., 13, 653–668.
Grebmeier, J.M., and L.W. Cooper, 1995. Influence of the St. Lawrence Island polynya upon the Bering Sea benthos. J. Geophys. Res., 100, 4439–4460.
Grebmeier, J.M., and L.W. Cooper, 2002. Benthic processes in the Bering Strait region of the Arctic: temporal/spatial variability and global change. Eos Trans. Amer. Geophys. Union, 83(4), Ocean Sciences Meeting Supplement, Abstract OS32M-01.
Grebmeier, J.M., and K.H. Dunton, 2000. Benthic processes in the Northern Bering/Chukchi Seas: status and global change. In: Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic, Huntington, H.P. (ed.), Marine Mammal Commission, Bethesda, Maryland, 61–71.
Hare, S.R., and N.J. Mantua, 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog. Oceanogr., 47, 103–146.
Harwood, L.A., P. Norton, B. Day, and P.A. Hall, 2002. The harvest of Beluga Whales in Canada's western Arctic: Hunter-based monitoring of size and composition of the catch. Arctic, 55(1), 10–20.
Hinckley, S., 1987. The reproductive biology of walleye pollock, Theragra chalcogramma, in the Bering Sea, with reference to spawning stock structure. Fish. Biol., 85, 481–498.
Hood, D.W., 1999. PROBES: Processes and Resources of the eastern Bering Sea shelf. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), University of Alaska Sea Grant, AK-SG-99-03, 697–712.
Hood D.W., and J.A. Calder, 1981. The eastern Bering Sea Shelf: oceanography and resources. University Wash. Press, Seattle, 520 pp.
Hood, D.W., and E.J. Kelley, 1974. Oceanography of the Bering Sea. Occasional Publication 2, Inst. of Marine Sci., Univ. of Alaska, Fairbanks, 623 pp.
Hunt, G.L., and N.M. Harrison, 1990. Foraging habitat and prey taken by Least Auklets at King Island, Alaska. Mar. Ecol. Prog. Ser., 65, 141–150.
Hunt, G.L., N.M. Harrison, and T. Cooney, 1990. Foraging of Least Auklets: The influence of hydrographic structure and prey abundance. Studies in Avian Biol. 14, 7–22.
Hunt, G.L., and P.J. Stabeno, 2002. Climate change and the control of energy flow in the southeastern Bering Sea. Prog. Oceanogr., 55, 5–22.
Hunt, G.L., C.L. Baduini, R.D. Brodeur, K.O. Coyle, N.B. Kachel, J.M. Napp, S.A. Salo, J.D. Schumacher, P.J. Stabeno, D.A. Stockwell, T.E. Whitledge, and S.I. Zeeman, 1999. The Bering Sea in 1998: The second consecutive year of extreme weather-forced anomalies. Eos Trans. AGU, 80(47), 561, 565–566.
Hunt, Jr., G.L., P. Stabeno, G. Walters, E. Sinclair, R. Brodeur, J.M. Napp, and N. Bond, 2002. Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep-Sea Res. II, Topical Studies in Oceanography, 49, 5821–5853.
Huntington, H.P., 2000. Traditional knowledge of the ecology of Belugas, Delphinapterus leucas, in Cook Inlet, Alaska. Mar. Fish. Rev., 62(3), 134–140.
Ianelli, J.N., T. Buckley, T. Honkalehto, N. Williamson, and G. Walters, 2001. Bering Sea-Aleutian Islands Walleye Pollock Assessment for 2002. In: Stock assessment and fishery evaluation report for the groundfish resources of the Bering Sea/Aleutian Islands regions, North Pac. Fish. Mgmt. Council, Anchorage, AK, section 1, 1–105.
Ikeda, M., 1990. Decadal oscillations of the air-ice-sea system in the northern hemisphere. Atmosphere- Ocean, 28, 106–139.
Incze, L.S., D.A. Armstrong, and S.L. Smith, 1987. Abundance of larval Tanner crabs (Chionoectes spp.) in relation to adult females and regional oceanography of the southeastern Bering Sea. Can. J. Fish. Aquat. Sci., 44, 1143–1156.
Incze, L.S., D. Hebert, N. Wolff, N. Oakey, and D. Dye, 2001. Changes in copepod distributions associated with increased turbulence from wind stress. Mar. Ecol. Prog. Ser., 213, 229–240.
Iverson, R.L., L.K. Coachman, R.T. Cooney, T.S. English, J.J. Goering, G.L. Hunt, Jr., M.C. Mc-Cauley, C.P. Mc Roy, W.S. Reesburgh, and T.E. Whitledge, 1979. Ecological significance of fronts in the southeastern Bering Sea. In: Ecological Processes in Coastal and Marine Systems, R.J. Livingston (ed.), Plenum Press, New York, 437–466.
Jin, Fei-Fei, M. Kimoto, and X. Wang, 2001. A model of decadal ocean-atmosphere interaction in the North Pacific basin. Geophys. Res. Lett., 28, 1531–1534.
Kachel, N.B., G. Hunt, S.A. Salo, J.D. Schumacher, P.J. Stabeno, and T.E. Whitledge, 2002. Characteristics of the Inner Front of the Southeastern Bering Sea. Deep Sea Res. II, Topical Studies in Oceanography, 49, 5889–5909.
Kendall, A.W., 2001. Specific gravity and vertical distribution of walleye pollock (Theragra chalcogramma) eggs. AFSC Processed Report 2001-01.
Khen, G.V., 1999. Hydrography of the western Bering Sea shelf water. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 161–176.
Kowalik, Z., and P. Stabeno (1999): Trapped motion around the Pribilof Islands in the Bering Sea. J. Geophys. Res., 104(C11), 25,667–25,684.
Kruse, G.H., 1998. Salmon run failures in 1997–1998: A link to anomalous ocean conditions? Alaska Fish. Res. Bull., 5, 55–63.
Labitzke, M., and H. van Loon, 1988. Associations between the 11-year solar cycle, the QBO and the atmosphere. I, The troposphere and stratosphere of the northern hemisphere in winter. J. Atmos. Terr. Phys., 50, 197–206.
Laevastu, T., and H.A. Larkins, 1981. Marine fisheries ecosystems: Its quantitative evaluation and management. Fishing News Books, Surrey, England, 162 pp.
Latif, M., and T. Barnett, 1994. Causes of decadal climate variability over the North Pacific and North America. Science, 266, 634–637.
Livingston, P.A., 1993. The importance of predation by groundfish, marine mammals, and birds on walleye pollock Theragra chalcogramma and Pacific herring Clupea pallasi in the eastern Bering Sea. Mar. Ecol. Prog. Ser., 102, 205–215.
Livingston, P.A., and R.D. Methot, 1998. Incorporation of predation into a population assessment model of eastern Bering Sea walleye pollock. In: Fishery Stock Assessment Models, Alaska Sea Grant College Program, AK-SG-98-01, 16 pp.
Livingston, P.A., L.L. Low, and R.J. Marasco, 1999. Eastern Bering Sea ecosystem trends. In: Large Marine Ecosystems of the Pacific Rim: Assessment, Sustainability, and Management, Q. Tang and K. Sherman (eds.), Blackwell Science, Boston, 140–162.
Longhurst, A.R., 1998. Ecological Geography of the Sea. Academic Press San Diego, 398 pp.
MacKenzie, B.R., T.J. Miller, S. Cyr, and W.C. Leggett, 1994. Evidence for a dome-shaped relationship between turbulence and larval fish ingestion rates. Limnol. Oceanogr., 39(8), 1790–1799.
McRoy, C.P., 1993. ISHTAR, the project: an overview of Inner Shelf Transfer And Recycling in the Bering and Chukchi seas. Cont. Shelf Res., 13, 473–480.
McRoy, C.P., 1999. Water over the Bridge: A summing up of the contributions of the ISHTAR Project in the Northern Bering and Chukchi Seas. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 687–697.
Macklin, S.A., V.I. Radchenko, S. Saitoh, and P.J. Stabeno, 2002. Variability in the Bering Sea ecosystem. Prog. Oceanogr., 55, 1–4.
McFarlane, G.A., J.R. King, and R.J. Beamish, 2000. Have there been recent changes to climate? Ask the fish. Prog. Oceanogr., 47, 147–169.
Megrey, B.A., and S. Hinckley, 2001. Effect of turbulence on feeding of larval fishes: A sensitivity analysis using an individual-based model. ICES J. Mar. Sci., 58, 1015–1029.
Merrick, R., 1995. The relationship of the foraging ecology of Stellar sea lions (Eumetopias jubatus) to their population decline in Alaska. Ph.D. Thesis, Univ. Washington, Seattle,Washington, USA, unpublished.
Merrick, R.L., 1997. Current and historical roles of apex predators in the Bering Sea ecosystem. J. Northwest Atlantic Fish. Sci., 22, 343–355.
Minobe, S., 1999. Resonance in bidecadal and pentadecadal climate oscillations over the North Pacific: Role in climate regime shifts. Geophys. Res. Lett., 26(7), 853–858.
Minobe, S., 2002. Interannual to interdecadal changes in the Bering Sea and concurrent 1998/99 changes over the North Pacific. Prog. Oceanogr., 55, 45–64.
Mizobata, K., S.I. Saitoh, A. Shiomoto, T. Miyamura, N. Shiga, K. Imai, M. Toratani, Y. Kajiwara, and K. Sasaoka, 2002. Bering Sea cyclonic and anticyclonic eddies observed during summer 2000 and 2001. Prog. Oceanogr., 55, 65–75.
Muench, R.D. (ed.), 1983. Marginal Ice Zones, J. Geophys. Res., 88, 2713–2966.
Muench, R.D., R.B. Tripp, and J.D. Cline, 1981. Circulation and hydrography of Norton Sound. In: The Eastern Bering Sea Shelf: Oceanography and Resources, Vol. 1, D.W. Hood and J.A. Calder (eds.), U.S. Government Printing Office, Washington D.C. (distributed by the University of Washington Press, Seattle), 77–93.
Napp, J.M., and G.L. Hunt, Jr., 2001. Anomalous conditions in the southeastern Bering Sea, 1997: linkages among climate, weather, ocean, and biology. Fish. Oceanogr., 10(1), 61–68.
Napp, J.M., A.W. Kendall, Jr., and J.D. Schumacher, 2000. A synthesis of biological and physical processes affecting the feeding of larval walleye pollock (Theragra chalcogramma) in the Eastern Bering Sea. Fish. Oceanogr., 9, 147–162.
Napp, J.M., C.T. Baier, K.O. Baier, R.D. Brodeur, N. Shiga, and K. Mier, 2002. Interannual and decadal variability in zooplankton communities of the southeast Bering Sea shelf. Deep-Sea Res. II, 49, 5991–6008.
Naumenko, N.I., 1996. Long-term fluctuations in the ichthyofauna of the Western Bering Sea. In: Ecology of the Bering Sea: A review of Russian literature, Mathiesen, O.A. and K.O. Coyle (eds.), University of Alaska Sea Grant Program Report No. 96-01, Fairbanks, AK, 143–158.
Niebauer, H.J., 1998. Variability in the Bering Sea ice cover as affected by a regime shift in the North Pacific in the period 1947–1996. J. Geophys. Res., 103, 27,717–27,737.
Niebauer, H.J., V. Alexander, and S.M. Henrichs, 1995. A time-series study of the spring bloom at the Bering Sea ice edge I: Physical processes, chlorophyll and nutrient chemistry. Cont. Shelf Res., 15, 1859–1878.
Nihoul, J.C.J., P.Adam, P. Brasseur, E. Deleersnijder, S. Djenidi, and J. Haus, 1993. Three-dimensional general circulation model of the Bering-Chukchi shelf. Cont. Shelf Res., 13, 509–542.
Noongwook, G., 2000. Native observations of local climate around St. Lawrence Island. In: Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic, Huntington, H.P. (ed.), Report of the Marine Mammal Commission Workshop, Marine Mammal Commission, Bethesda, Maryland, 21–24.
NRC Report, 1996. The Bering Sea ecosystem. National Academy Press, 307 pp.
NRC (2003): The decline of the Steller sea lion in Alaskan waters: Untangling Food Webs and Fishing Nets. National Academy Press, Washington, D.C., 204 pp.
Ohtani, K., and T. Azumaya, 1995. Influence of interannual changes in ocean conditions on the abundance of walleye pollock (Theragra chalcogramma) in the eastern Bering Sea. In: Climate Change and northern fish populations, R.J. Beamish (ed.), Can. Spec. Pub. Fish. Aquat. Sci., 121, 87–95.
Olla, B.L., and M.W. Davis, 1990. Effects of physical factors on the vertical distribution of larval walleye pollock Theragra chalcogramma under controlled conditions. Mar. Ecol. Prog. Ser., 63, 105–112.
Olson, M.B., and S.L. Strom, 2002. Phytoplankton growth, microzooplankton herbivory and community structure in the southeast Bering Sea: insight into the formation and temporal persistence of an Emiliania huxleyi bloom. Deep-Sea Res. II, Topical Studies in Oceanography, 49, 5969–5990.
Overland, J.E., 1981. Marine climatology of the Bering Sea. In: The Eastern Bering Sea Shelf: Oceanography and Resources, Vol. 1, D.W. Hood and J.A. Calder (eds.), U.S. Government Printing Office, Washington D.C. (distributed by the University of Washington Press, Seattle), 15–30.
Overland, J.E., and C.H. Pease, 1982. Cyclone climatology of the Bering Sea and its relation to sea ice extent. Mon. Weather Rev., 110, 5–13.
Overland, J.E., and A.T. Roach, 1987. Northward flow in the Bering and Chukchi Seas. J. Geophys. Res., 92(C7), 7097–7105.
Overland, J.E., M.C. Spillane, H.E. Hurlburt, and A.J. Wallcraft, 1994. A numerical study of the circulation of the Bering Sea basin and exchange with the North Pacific Ocean. J. Phys. Oceanogr., 24, 736–758.
Overland, J.E., J.M. Adams, and N.A. Bond, 1999a. Decadal variability of the Aleutian Low and its relation to high-latitude circulation. J. Climate, 12, 1542–1548.
Overland, J.E, S.A. Salo, L.H. Kantha, and C.A. Clayson, 1999b. Thermal stratification and mixing on the Bering Sea shelf. In: Dynamics of The Bering Sea, Loughlin, T.R., and K. Ohtani (eds.), Alaska Sea Grant College Program, Fairbanks, AK, 129–146.
Parrish, R.H., F.B. Schwing, and R. Mendelssohn, 2000. Mid-latitude wind stress: the energy source for climate shifts in the North Pacific Ocean. Fish. Oceanogr., 9, 224–238.
Peterson, W.T., and F.B. Schwing, 2003. A new climate regime in northeast Pacific ecosystems. Geophys. Res. Lett., 30, OCE 6-1 to 6-4.
Pungowiyi, C., 2000. Native observations of change in the marine environment of the Bering Sea. In: Impacts of Changes in Sea Ice and Other Environmental Parameters in the Arctic, Huntington, H.P. (ed.), Report of the Marine Mammal Commission Workshop, Marine Mammal Commission, Bethesda, Maryland, 18–20.
Radchenko, V.I., G.V. Khen, and A.M. Slabinsky, 2001. Cooling in the western Bering Sea in 1999: quick propagation of La Ni˜na signal or compensatory processes effect? Prog. Oceanogr., 49, 407–422.
Reed, R.K., 1995. On the variable subsurface environment of fish stocks in the Bering Sea. Fish. Oceanogr., 4, 317–323.
Reed, R.K., 1998. Confirmation of a convoluted flow over the Eastern Bering Sea Shelf. Cont. Shelf Res., 18, 99–103.
Reed, R.K., and P.J. Stabeno, 1999. The Aleutian North Slope Current. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), University of Alaska Sea Grant, AK-SG-99-03, 177–192.
Rosenkranz, G.E., A.V. Tyler, and G.H. Kruse, 2001. Effects of water temperature and wind on recruitment of Tanner crabs in Bristol Bay, Alaska. Fish. Oceanogr., 10, 1–12.
Royer, T.C., 1998. Coastal processes in the northern North Pacific. In: The Sea: Vol. 11—The Global Coastal Ocean: Regional Studies and Synthesis, John Wiley & Sons, Inc., New York, NY, 395–414.
Royer, T.C., C.E. Gorsch, and L.A. Mysak, 2001. Interdecadal variability of Northeast Pacific coastal freshwater and its implications on biological productivity. Prog. Oceanogr., 49, 95–111.
Sambrotto R.N., H.J. Niebauer, J.J. Goering, and R.L. Iverson, 1986. Relationships among vertical mixing nitrate uptake and phytoplankton growth during the spring bloom in the southeast Bering Sea middle shelf. Cont. Shelf Res., 5, 161–198.
Schumacher, J.D., and V. Alexander, 1999. Variability and role of the physical environment in the Bering Sea ecosystem. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 147–160.
Schumacher, J.D., and T.H. Kinder, 1983, Low frequency currents over the Bering Sea Shelf. J. Phys. Oceanogr., 13, 607–623.
Schumacher, J.D., and A.W. Kendall, Jr., 1995. Fisheries Oceanography: Walleye Pollock in Alaskan Waters. Rev. Geophys., Suppl., 33, 1153–1163.
Schumacher, J.D., and R.K. Reed, 1992. Characteristics of currents near the continental slope of the eastern Bering Sea. J. Geophys. Res., 97, 9423–9433.
Schumacher, J.D., and P. J. Stabeno, 1994. Ubiquitous eddies of the Eastern Bering Sea and their coincidence with concentrations of larval pollock. Fish. Oceanogr., 3, 182–190.
Schumacher, J.D., and P. J. Stabeno, 1998. Continental shelf of the Bering Sea. In: The Sea: Vol. 11—The Global Coastal Ocean: Regional Studies and Synthesis, John Wiley & Sons, Inc., New York, NY, 789–822.
Schumacher, J.D., K. Aagaard, C.H. Pease, and R.B. Tripp, 1983. Effects of a shelf polynya on flow and water properties in the northern Bering Sea. J. Geophys. Res., 88, 2723–2732.
Schumacher, J.D., N.A. Bond, R.D. Brodeur, P.A. Livingston, J.M. Napp, and P.J. Stabeno, 2003. Climate Change in the Southeastern Bering Sea and Some Consequences for Biota. In: Large Marine Ecosystems of the World: Trends in Exploitation, Protection, and Research, G. Hempel, and K. Sherman (eds.), Amsterdam: Elsevier Science, 17–40.
Shuert, P.G., and J.J. Walsh, 1993. A coupled physical-biological model of the Bering-Chukchi seas. Cont. Shelf Res., 13, 543–573.
Sirenko, B.I., and V.M. Koltun, 1992. Characteristics of the benthic biocenoses of the Chukchi and Bering Seas. In: Results of the third joint U.S.-U.S.S.R. Bering and Chukchi Sea Expedition (BERPAC), P.A. Nagel (ed.), U.S. Fish and Wildlife Service, Washington, DC, 251–261.
Smith, S.L., and J. Vidal, 1986. Variations in the distribution, abundance, and development of copepods in the southeastern Bering Sea in 1980 and 1981. Cont. Shelf Res., 13, 215–240.
Springer, A.M., 1999. Summary, conclusions and recommendations. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 777–800.
Springer, A.M., and G.V. Byrd, 1989. Seabird dependence on walleye pollock in the Eastern Bering Sea. In: Proceedings of the International Symposium on the Biology and Management of Walleye Pollock, Alaska Sea Grant, AK-SG-89-01, 667–677.
Springer, A.M., and C.P. McRoy, 1993. The paradox of pelagic food webs in the northern Bering Sea—III. Patterns of primary production. Cont. Shelf Res., 13, 575–600.
Springer, A.M., E.C. Murphy, D.G. Rosenau, C.P. McRoy, and B.A. Cooper, 1987. The paradox of pelagic food webs in the northern Bering Sea—I. Seabird food habits. Cont. Shelf Res., 7(8), 895–911.
Springer, A.M., C.P. McRoy, and K.R. Turco, 1989. The paradox of pelagic food webs in the northern Bering Sea—II. Zooplankton communities. Cont. Shelf Res., 9, 359–386.
Springer, A.M., C.P. McRoy, and M.V. Flint, 1996. The Bering Sea Green Belt: shelf-edge processes and ecosystem production. Fish. Oceanogr., 5, 205–223.
Springer, A.M., and D.G. Roseneau, 1985. Copepod-based food webs: auklets and oceanography in the Bering Sea. Mar. Ecol. Prog. Ser., 21, 229–237.
Stabeno, P.J., and G.L. Hunt, 2002. Overview of the Inner Front and Southeast Bering Sea Carrying Capacity Programs. Deep-Sea Res. II: Topical Studies in Oceanography, 49, 6157–6168.
Stabeno, P.J., and J.E. Overland, 2001. Bering Sea shifts toward an earlier spring transition. Eos Trans. AGU, 29, 317, 321.
Stabeno, P.J., R.K. Reed, and J.D. Schumacher, 1995: The Alaska Coastal Current: Continuity of transport and forcing. J. Geophys. Res., 100, 2477–2485.
Stabeno, P.J., J.D. Schumacher, R.F. Davis, and J.M. Napp, 1998. Under-ice observations of water column temperature, salinity and spring phytoplankton dynamics: Eastern Bering Sea shelf, 1995. J. Mar. Res., 56, 239–255.
Stabeno, P.J., J.D. Schumacher, and K. Ohtani, 1999. The physical oceanography of the Bering Sea. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 1–28.
Stabeno, P.J., N.A. Bond, N.B. Kachel, S.A. Salo, and J.D. Schumacher, 2001. On the temporal variability of the physical environment over the southeastern Bering Sea. Fish. Oceanogr., 10(1), 81–98.
Stabeno, P.J., N.B. Kachel,M. Sullivan, and T.E. Whitledge, 2002. Variability along the 70-m isobath of the southeast Bering Sea. Deep-Sea Res. II: Topical Studies in Oceanography, 49, 5931–5943.
Stabeno, P.J., D.K. Kachel, N.A. Kachel, and M.A. Sullivan, 2004. Observations from moorings in the Aleutian passes: Temperature, salinity, and transport. Fish. Oceanogr., 14(suppl. 1) 39–54.
Stenseth, N.C., A. Mysterud, G. Ottersen, J.W. Hurrell, K.-S. Chan, and M. Lima, 2002. Ecological effects of climate fluctuations. Science, 297, 1292–1296.
Stockwell, D.A., T.E. Whitledge, S.I. Zeeman, K.O. Coyle, J.M. Napp, R.D. Broduer, A.I. Pinchuk, and G.L. Hunt, Jr., 2001. Anomalous conditions in the southeastern Bering Sea, 1997: Nutrients, phytoplankton, and zooplankton. Fish. Oceanogr., 10, 99–116.
Sugimoto, T., and Tadokoro, K., 1997. Interannual–interdecadal variations in zooplankton biomass, chlorophyll concentration and physical environment in the subarctic Pacific and Bering Sea. Fish. Oceanogr., 6, 74–93.
Takenouti, Y., and K. Ohtani, 1974. Currents and water masses in the Bering Sea: A review of Japanese work. In: Oceanography of the Bering Sea, D.W. Hood and E.J. Kelley (eds.), Occasional Publication Number 2, Institute of Marine Science, University of Alaska, Fairbanks, 39–57.
Tourangeau, S., J.A. Runge, 1991. Reproduction of Calanus glacialis under ice in spring in southeastern Hudson Bay, Canada. Mar. Biol., 108, 227–233.
Trites, A.W., P.A. Livingston, M.C. Vasconcellos, S. Mackinson, A.M. Springer, and D. Pauly, 1999. Ecosystem change and the decline of marine mammals in the Eastern Bering Sea: testing the ecosystem shift and commercial whaling hypothesis. Fisheries Centre Reports, 7(1), 98 pp.
Tsyban, A.V., 1999. The BERPAC Project: development and overview of the ecological investigation in the Bering and Chukchi Seas. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), University of Alaska Sea Grant, AK-SG-99-03, 697–712.
Tynan, C.T., 1998. Coherence between whale distributions, chlorophyll concentration, and oceanographic conditions on the southeast Bering Sea shelf during a coccolithophore bloom, July– August, 1997. Eos Trans. AGU, 79, 127.
Vance, T.C., J.D. Schumacher, P.J. Stabeno, C.T. Baier, T. Wyllie-Echeverria, C.T. Tynan, R.D. Brodeur, J.M. Napp, K.O. Coyle, M.B. Decker, G.L. Hunt, Jr., D. Stockwell, T.E. Whitledge, M. Jump, and S. Zeeman, 1998. Aquamarine waters recorded for first time in eastern Bering Sea. Eos Trans. AGU, 79, 121, 126.
Verkhunov, A.V., 1994. Thermohaline characteristics of the shelf front in the western part of the Bering Sea, Oceanology, 34, 356–369.
Verkhunov, A.V., and Y.Y. Tkachenko, 1992. Recent observations of variability in the Western Bering Sea Current system. J. Geophys. Res., 97, 14,369–14,376.
Walsh, J.J., and C.P. McRoy, 1986. Ecosystem analysis in the southeastern Bering Sea. Cont. Shelf Res., 5, 259–288.
Walsh, J.J., C.P. McRoy, L.K. Coachman, J.J. Georing, J.J. Nihoul, T.E. Whitledge, T.H. Blackburn, P.L. Parker, C.D. Wirick, P.G. Shuert, J.M. Grebmeier, A.M. Springer, R.D. Tripp, D.A. Hansell, S. Djenidi, E. Deleersnijder, K. Henriksen, B.A., Lund, P. Andersen, F.E. Muller-Karger, and K. Dean, 1989. Carbon and nitrogen recycling with the Bering/Chukchi Seas: source regions for organic matter effecting AOU demands of the Arctic Ocean. Prog. Oceanogr., 22, 277–359.
Walters, C.J., V. Christensen, D. Pauly, and J. Kitchell, 1999. ECOSIM II: Representation of trophic interaction effects in a population dynamics model. Ecosystems, 3, 70–83.
Wespestad, V.G., L.W. Fritz, W.J. Ingraham, Jr., and B.A. Megrey, 2000. On relationships between cannibalism, climate variability, physical transport and recruitment success of Bering Sea walleye pollock, Theragra chalcogramma. ICES J. Mar. Sci., 57, 272–278.
Wilderbuer, T.K., A.B. Hollowed, W.J. Ingraham, Jr., P.D. Spencer, M.E. Conners, N.A. Bond, and G.E. Walters, 2002. Flatfish recruitment response to decadal climate variability and ocean conditions in the eastern Bering Sea. Prog. Oceanogr., 55, 235–247.
Wooster,W.S., and A.B. Hollowed, 1995. Decadal-scale variations in the eastern subarctic Pacific. I. Winter ocean conditions. Can. Spec. Publ. Fish. Aquat. Sci., 121, 81–85.
Wyllie-Echeverria, T., 1995. Sea-ice conditions and the distribution of walleye pollock and the Bering and Chukchi Sea shelf. In: Climate Change and Northern Fish Populations, R.J. Beamish (ed.), Can. Spec. Publ. Fish. Aquat. Sci., 121, 87–95.
Wyllie-Echeverria, T., and K. Ohtani, 1999. Seasonal sea ice variability and the Bering Sea ecosystem. In: Dynamics of the Bering Sea: A Summary of Physical, Chemical, and Biological Characteristics, and a Synopsis of Research on the Bering Sea, T.R. Loughlin and K. Ohtani (eds.), North Pacific Marine Science Organization (PICES), Univ. of Alaska Sea Grant, AK-SG-99-03, 435–452.
Zheng, J., G.H. Kruse, and D.A. Ackley, 2001. Spatial distribution and recruitment patterns of Snow crabs in the eastern Bering Sea. In: Spatial process and management of marine populations, Alaska Sea Grant College Program, AK-SG-01-02, 233–255.
Return to Abstract